Cita: Scofield HN, Mattila HR (2015) Honey Bee Trabajadores Que son el polen Destacó como larvas se empobrecen Las recolectores y Waggle bailarines como adultos. PLoS ONE 10 (4): e0121731. doi: 10.1371 / journal.pone.0121731
Editor Académico: Olav Rueppell, Universidad de Carolina del Norte, Greensboro, Estados Unidos
Recibido: 26 de junio 2014; Aceptado: 18 de febrero 2015, Publicado: 08 de abril 2015
Abstracto
Los efectos negativos sobre la conducta de los adultos de la desnutrición de menores están bien documentados en los vertebrados, pero relativamente poco conocidos en los invertebrados. Hemos examinado los efectos del estrés nutricional de larvas en el comportamiento de forrajeo y la contratación de un importante económicamente invertebrado modelo, la abeja de la miel ( Apis mellifera ).
 |
| Fig 1. Peso, longevidad, y la actividad de forrajeo de los trabajadores adultos se redujo cuando el acceso al polen se limitó durante el desarrollo larval. |
El polen, que suministra los nutrientes esenciales a los trabajadores en desarrollo, puede llegar a ser limitada en las colonias debido a dearths estacionales, la pérdida de hábitat de alimentación, o el manejo intensivo. Sin embargo, las consecuencias funcionales de ser criados por compañeros de nido de polen estresado siguen sin estar claros, a pesar de la creciente preocupación de que la mala nutrición interactúa con otros factores de estrés para exacerbar declive colonia.
Manipulamos acceso enfermera abeja al polen y luego evaluaron diferencias en el peso, la longevidad, la actividad de forrajeo, y el comportamiento de la abeja-danza de los trabajadores que crían (que eran co-promovidos como adultos). Polen estrés durante el desarrollo larval tuvo trascendentales efectos físicos y de comportamiento en los trabajadores adultos.
Trabajadores criados en colonias de polen estresadas eran más ligeras y más corta duración que compañeros de nido criados con un acceso adecuado al polen. Proporcionalmente menos trabajadores estresados se observaron zonas de alimentación y los que lo hicieron forraje comenzaron forrajeo antes, forrajearon por menos días, y tenían más probabilidades de morir después de un solo día de forrajeo.Trabajadores de polen estresadas eran también menos propensos a menear la danza que sus contrapartes no acentuadas y, si bailaban, la información que transmiten sobre la ubicación de la comida era menos precisa. Estos déficits de rendimiento pueden escalar si evita la limitación del polen a largo plazo destacaron recolectores de proporcionar suficiente para el desarrollo de los trabajadores.
Por otra parte, los efectos de la escasez de polen breves informaron aquí reflejan los efectos de otros factores de estrés ambientales que limitan el acceso de los trabajadores a los nutrientes, lo que sugiere la posibilidad de que su interacción sinérgica.
Las abejas de miel a menudo experimentan el nivel de estrés que hemos creado, por tanto, nuestros resultados ponen de relieve la importancia de una nutrición adecuada para apoyar el desempeño del trabajador y su potencial contribución a la productividad de la colonia y los servicios de polinización de calidad.
Introducción
El impacto negativo del estrés nutricional de desarrollo en función de los adultos ha sido documentado en una amplia gama de vertebrados (revisado por [ 1 - 7 ]). Las consecuencias funcionales de estrés comida temprana son diversas, incluyendo el aprendizaje y la canción producción alterada en los pájaros cantores adultos [ 8 ], la mala respuesta de las ranas al estanque secado [ 9 ], y metabólica de aparición tardía y trastornos del desarrollo neurológico en los seres humanos [ 10 - 13 ]. Por el contrario, nuestra comprensión de los efectos de la desnutrición en el desarrollo en el rendimiento de los adultos en los invertebrados es relativamente delgado. Los estudios con insectos modelo muestran que el acceso insuficiente a los alimentos durante el desarrollo larvario puede retrasar "ritmo de vida" [ 14 ] al disminuir el crecimiento y el inicio de la reproducción como una estrategia adaptativa para soportar la escasez de alimentos (por ejemplo, las moscas de la fruta [ 15 , 16 ]; mariquita escarabajos [17 ]; mosquitos [ 18 ]), que pueden resultar en un menor peso corporal adulto e imponer costos reproductivos de las personas [ 19 - 22 ]. Pocos estudios de insectos han examinado los efectos del estrés alimento larval sobre el comportamiento adulto no reproductiva, aunque se ha demostrado que disminuye el metabolismo de vuelo y la defensa del territorio de las mariposas [ 23 , 24 ] y alterar forrajeo exploratoria, la capacidad de aprendizaje y la memoria en la fruta adulta moscas [ 25 , 26 ].
El objetivo de este estudio es evaluar las consecuencias del estrés alimento larval para adultos de otro modelo importante de invertebrados-la abeja de la miel ( Apis mellifera L.). Estrategia de miel de abeja para lidiar con el estrés de alimentos se complica por eusocialidad, donde "ritmo de vida" ajustes son desafiados por la enorme energía y laborales demandas de crianza colectiva cría, forrajeo, y el mantenimiento del nido. La necesidad de entender los efectos potenciales de estrés nutricional de las abejas melíferas ha vuelto urgente en los últimos años.Las abejas de miel polinizan casi la mitad de los cultivos que se cultivan en todo el mundo para su uso por los seres humanos [ 27 ] y son factores clave para el estimado € 153 mil millones (~ $ 211 mil millones de dólares) del valor anual total de la polinización por insectos [ 28 ]. Sin embargo, el papel de miel de abeja en la producción mundial de alimentos ha sido amenazada en los últimos años por la inusualmente alta mortalidad de colonias manejadas [ 29 - 38 ], junto con el número de colonias en declive durante las décadas anteriores [ 36 , 39 ]. Consenso creciente de que los efectos e interacciones de un conjunto de múltiples factores de estrés, incluyendo parásitos, patógenos, pesticidas, baja diversidad genética y la mala alimentación, está causando o agravando estas pérdidas [ 39 - 41 ]. El estrés nutricional es una preocupación particular porque puede actuar sinérgicamente con otros factores de estrés ambientales [ 42 , 43 ] y se presume agravándose en zonas con hábitat disminución de forrajeo, donde las pérdidas de colonias son el mayor [ 44 ].
La principal fuente de estrés nutricional en las colonias es el acceso inadecuado al polen, que proporciona las proteínas esenciales, lípidos, vitaminas y minerales que son necesarios para el desarrollo larvario y la función adulta [ 45 , 46 ]. En consecuencia, la disponibilidad de polen está estrechamente relacionado con el número de abejas de la miel que una colonia puede trasero [ 47 - 50 ]. Colonias experimentan rutinariamente escasez de polen de temporada cuando las colonias agotan tiendas antes más polen puede ser recogida [ 51 - 53 ], como a menudo ocurre durante breves periodos de mal tiempo [ 49 , 54 - 56 ] o como un subproducto no deseado de las prácticas de gestión comercial que ponen colonias en una intensa competencia por las fuentes de polen que pueden carecer de la diversidad, ser poco nutritiva, o flores con poca frecuencia [ 57 - 63 ]. En respuesta a la escasez de polen, las colonias tienden a ajustar el número de larvas que puedan trasera a través de la utilización de sus propias reservas corporales para apoyar al empollar [ 64 ] y por canibalizar las larvas jóvenes e invertir nutrientes recuperados en las larvas mayores [ 49 , 56 , 65 ] . A pesar de estas medidas de protección, los adultos de tamaño insuficiente y agotados de nutrientes a veces son criados cuando el acceso al polen es limitada [ 66 - 68 ].
Estudios previos que han probado los efectos de la manipulación de la nutrición de las larvas en los adultos han utilizado dietas artificiales, larvas criadas a mano, los adultos enjaulados, o alguna combinación de los mismos (por ejemplo, [ 69 - 75 ], pero ver [ 76 ] para una excepción). Sin embargo, la evaluación más realista de las consecuencias de la desnutrición trabajador requiere efectos en un contexto social natural de instrucción. Nuestro estudio, que se repitió tres veces, se reúne este criterio. Debido a que nuestros trabajadores focales vivían en un medio social normal, hemos sido capaces de examinar los efectos de ser criados en colonias de polen subrayado en dos tareas complejas que se realizan posteriormente en la vida de los adultos trabajadores de forrajeo y la contratación. Creamos cohortes de trabajadores nutricionalmente tónicas y átonas que fueron criados en colonias por enfermeras nestmate con acceso a cualquiera de las tiendas de polen limitados o adecuadas.Trabajadores Focales se pesaron en la emergencia de adultos y el entonces fomentaron co como adultos en las mismas colonias de acogida, donde se ha monitorizado su longevidad, la actividad de forrajeo, y el comportamiento de la abeja-dance. Hemos encontrado que el estrés polen durante el desarrollo larvario tenía de largo alcance efectos negativos en el desempeño de tareas de los adultos en el futuro. Fundamentalmente, los déficits de rendimiento extenderse a zonas de alimentación y selección de personal, que son las tareas más importantes que las abejas realizan como abastecedores de sus colonias y como polinizadores de cultivos humanos cultivados.
Materiales y METODOS
La manipulación de la disponibilidad del polen durante el desarrollo larval
En tres ensayos independientes llevados a cabo durante dos años, se comparó el peso, la longevidad, la actividad de forrajeo, y el rendimiento de la abeja-dance de los trabajadores de abejas que fueron criados como larvas en condiciones de suministro de polen sea limitada o abundante, pero compartimos un entorno social común como adultos (2012: Ensayo 1; 2013: ensayos 2 y 3; espectáculo de danza se examinó en el ensayo 3 solamente).
Para manipular las condiciones de desarrollo para los trabajadores de centros de coordinación en cada ensayo, nos dividimos las colonias de origen en tres subunidades por colonia, con el objetivo de crear una subunidad polen limitado que agotar su suministro de polen durante el desarrollo de las larvas de coordinación y otros dos subunidades que se encontraban en abundancia suministrada con el polen para que las larvas focal se aprovisiona adecuadamente durante al empollar (cada uno era un tipo diferente de control, véase más adelante).Subunidades de una colonia se dejaron ya sea en la colmena original o transferidos a una de las dos cajas de colmena 5-frame. Todas las subunidades de colonias tenían larvas hacia atrás, pero variaban en la cantidad de polen que tenían que hacerlo. Una subunidad de cinco marco fue polen limitado: tenía 1-2 marcos de cría que contenía <50 cm="" font="" nbsp="">2 de peine polen lleno en total; polen adicional se raspó fuera de los marcos y todos los marcos restantes contenía miel solamente (pruebas anteriores de nuestro método demostraron que las colonias con más polen no agotan de forma fiable sus tiendas durante la cría del pollo). Subunidades de polen limitada también se les impidió la recogida de más polen, como ocurriría durante un período de tiempo desfavorable, mediante el cribado de sus entradas y colocarlos en una incubadora fresco para la duración del desarrollo de las larvas focal (10-12 ° C; VWR incubador de baja temperatura, Radnor, PA, EE.UU.). Estamos seguros de que estas subunidades de colonias eran polen limitada debido a que sus tiendas de polen siempre habían desaparecido a finales de desarrollo de las larvas de los trabajadores de atención, en contraste con los controles suministrados en abundancia (véase más adelante). Por otra parte, de las colonias 23 de origen que repartirá en todos los ensayos, las subunidades de polen limitada de sólo 7 colonias producen suficiente cría para ser incluidos en el estudio porque las colonias restantes canibalizado sus crías, presumiblemente en respuesta a la escasez de polen que nuestra manipulación creado [ 49 , 56 , 66 ] (ensayos 1 y 3 utilizaron tres colonias de origen cada uno, el ensayo 2 utiliza una colonia de origen). Un segundo conjunto de subunidades de colonias de cinco marco a juego estas subunidades de polen limitada se ensambla a partir de las colonias de origen y encierra de manera similar, excepto que uno de los marcos de alimentos en cada era al menos 90% lleno de polen en un lado (> 775 cm 2 de peine polen lleno, enfrentado hacia la cría), además de polen en cualquiera de los otros marcos (por lo tanto, los controles con abundante polen confinado). El tercer conjunto de subunidades (el resto de cada colonia de origen en la colmena el original) se le dio un marco de polen igualmente bien surtida y quedó confinado para que los trabajadores podían recoger más polen del medio ambiente (por lo tanto, los controles no confinados con abundante polen). Porque siempre había polen queda en subunidades de control al final del desarrollo de las larvas de los trabajadores focales (> 50% de su marco polen original), que confía en que las enfermeras que crían los trabajadores focales tenían acceso adecuado al polen durante el aprovisionamiento de las larvas. Cuadros de cría Las subunidades de colonias contenían crías de todas las edades, pero cuadros de cría fueron cuidadosamente distribuido entre subunidades de modo que cada uno tenía un marco con huevos y / o larvas jóvenes (<2 as="" d="" de="" edad="" font="" nbsp="">No nos etiquetamos adultos emergentes hasta por lo menos 17 días después se ensamblan subunidades, que aseguraron que los trabajadores focales experimentaron condiciones de tratamiento para la mayoría, si no todos, de su desarrollo larvario. Reina de cada colonia fuente fue dejado en la subunidad colmena originales; las nuevas colmenas recibieron dos señuelos impregnados con compuestos que se producen por las reinas para señalar su presencia, que suprime efectivamente reina de cría [77 ] (fueron reemplazados señuelos cada 2 días, las colonias que recibieron señuelos no mostraron signos de trabajador ponedoras o el reemplazo de la reina; Boost Bee, Phero Tech, Victoria, BC, Canadá). Cada subunidad colonia tenía 3-5 marcos trabajador cubierto sacudido en ellos, dependiendo del tamaño de la colonia fuente original. En el ensayo 1, no había trabajadores de control de confinados y no se utilizaron los señuelos de la reina; nos informan de este ensayo, sin embargo, debido a que sus hallazgos son similares a los de los otros dos ensayos para que todos los tratamientos de control estaban en su lugar.
Después de larvas focal había sido alimentada por sus compañeros de nido y sellado en las células para pupar, trabajadores adultos fueron retirados de todos los cuadros de cría, que se transfirieron luego a una incubadora caliente (35 ° C) cuando los trabajadores completaron su desarrollo. Marcos se comprobaron todos los días y, una vez adultos focales empezaron a emerger de celdas selladas, se pesaron individualmente en una balanza analítica (a 0,0001 g; AB104-S, Mettler Toledo, Columbus, OH, EE.UU.) y luego etiquetados en sus thoraxes con discos únicamente identificables numerados de plástico (BioQuip, CA, USA y Chr. Graze, Weinstadt, Alemania) antes de la introducción en la observación de la colmena de su juicio en el mismo día de la emergencia. Trabajadores Focales surgieron durante un período de 7 días en el ensayo 1 y un período de 4 días en los ensayos 2 y 3; entre los ensayos, el número medio de trabajadores focales introducidas por colonia fuente fue similar entre tratamientos (122 ± 30 trabajadores por colonia polen limitado subunidad; 121 ± 24 trabajadores por el control confinado con abundante polen; 128 ± 18 trabajadores por el control no confinado con abundante polen; F 2,15 = 0,2, P = 0,98). Los observadores no estaban al tanto del tratamiento al que pertenecían los trabajadores durante la recolección de datos subsiguiente. Para cada ensayo, una colonia del anfitrión se instaló en una colmena de observación queenright, dos-marco, que fue creado en el interior de una casa colmena en el Colegio Arboretum Wellesley (Wellesley, MA, EE.UU.). Todas las colonias de origen y de acogida (y sus reinas, que eran de ascendencia mixta y menos de un año de edad) se mantiene antes de cada ensayo en los apiarios de investigación Wellesley College después de haber sido comprado a proveedores de abejas locales (2012: Beehavin 'Colmenares, Smithfield, RI, EE.UU., 2013: Merrimack Valley Colmenares, Billerica, MA, EE.UU.). Los trabajadores de cada colmena de observación podían alimentarse al aire libre caminando a través de un tubo que conecta la colmena a una abertura en una pared del edificio. Se realizaron todos los ensayos durante los meses de verano (junio-agosto) y las colonias anfitrionas mantienen reservas de miel y polen por la duración de ese período (la colonia del anfitrión en el ensayo 3 corrió bajo en comida en un punto, por lo que uno de sus cuadros fue sustituido por un marco que contiene el polen y la miel). Todas las colonias de origen y de destino eran floreciente y aparecieron libres de enfermedad antes del inicio de cada ensayo.
La evaluación de la longevidad y de forrajeo desempeño de los trabajadores focales
La longevidad de los trabajadores focales se determinó mediante la comprobación de las colmenas de observación dos veces al día después de las primeras etiquetado trabajadores se introdujeron en ellos hasta que no se encontraron más trabajadores. Hemos llevado a cabo la mayoría de los controles en la mañana temprano y por la noche, cuando la actividad de forrajeo fue mínima y tuvimos la mayor oportunidad de ver a todos los trabajadores etiquetados. Registros de actividad (véase más adelante) de forrajeo se combinaron con datos colmena de verificación para mejorar la precisión de las estimaciones de longevidad. Hemos tomado nota de las identidades de los trabajadores etiquetados que no se veían en su colmena de observación 24 horas después de su introducción a la misma; sólo aquellos trabajadores que estaban presentes durante más de 24 horas se incluyeron en las comparaciones de tratamiento significa para el peso y desempeño de los trabajadores.
Actividad de forrajeo fue evaluada mediante el control de la entrada de cada colmena para los trabajadores de centros de coordinación en un período de 2 horas 09 a.m.-5 p.m. cada día.Una pista cubierta de plexiglás (26 x 10 x 3 cm) se adjuntó a cada entrada para facilitar la observación de los cazadores-recolectores etiquetados. Una serie de deflectores escalonados aumentó la distancia que entran y salen de las abejas tenían que viajar, lo que retrasó la mayoría de los trabajadores lo suficiente como para permitir su identificación. Las observaciones de forrajeo comenzaron cuando controles frecuentes revelaron que los trabajadores focales estuvieron presentes en su entrada de la colmena (día 7 en los ensayos 1 y 2 y el día 6 en el ensayo 3) y terminaron cuando el número de trabajadores focales había disminuido hasta el punto de que rara vez se observa (día 50 para el ensayo 1; 26 días para el juicio 2; 52 días para prueba de 3). La participación en búsqueda de alimento, la edad de inicio de la búsqueda de alimento, y el número de días observados forrajeo se compararon entre los grupos de tratamiento para cada ensayo.
Evaluar el desempeño de la abeja-dance de los trabajadores focales
El efecto de ser criados en colonias de polen limitada durante el desarrollo larvario en el comportamiento de la abeja-baile para adultos se estimó para los trabajadores de centros de coordinación en la prueba 3. Cada colmena de observación tuvo un solo "aquí bajo mi estrado, el baile área de peine junto a la entrada, donde bailan los forrajeros -porque una derivación obligado a todos los cazadores-recolectores que entran a entrar en el mismo lado de la estructura inferior. La pista de baile fue filmado durante 1-2 horas por día 09 a.m.-5 p.m. como el clima y recolector actividad permitida (con iluminación fluorescente de techo difuso), comenzando cuando los trabajadores focales se observaron por primera baile durante los controles frecuentes de la pista de baile (cuando el los trabajadores más jóvenes tenían 12 días de edad) y que termina cuando sólo una danza fue realizada por hora durante el día final de la grabación (cuando los trabajadores de más edad eran 45 días de edad; 41 horas de vídeo en total; Sony Handycam DCR-HC63, Tokio, Japón) . Recolectoras Focales realizan bailes waggle fueron identificados en cada cinta de vídeo por un observador, que más tarde se analizó cuadro por cuadro (un cuadro = 1/30 s) con Final Cut Express 4.0.1 (Apple Inc., Cupertino, CA, EE.UU.) para calcular métricas de baile. Una sola Danza de la abeja incluye todas las corridas waggle que fueron realizados por un recolector a su regreso a la colmena antes de que sea de izquierda a alimentarse de nuevo o se mueve más en la colmena. Un solo de danza por un recolector a menudo consiste en múltiples episodios de baile interrumpidas por pausas para cambiar la ubicación o para transferir alimentos a otros trabajadores o en las células; carreras totales waggle se suman para todos los segmentos de un baile que fueron interrumpidos por dichas pausas. Varios aspectos de la actividad de la abeja-dance se compararon entre los grupos de tratamiento para las danzas realizadas por los trabajadores de coordinación, ya que se alimentaban libremente a las fuentes de alimentos desconocidos: la participación en el baile, el número total de número de días observados por el baile bailarín, bailes totales por bailarín, de carreras waggle por bailarín, y se ejecuta promedio por la danza de cada bailarín. Duración Waggle a ejecutar (como sustituto de la distancia a las fuentes de alimentos) también se comparó entre un subconjunto de los trabajadores para determinar si el estrés polen desarrollo afectó a la distancia a la que los trabajadores se alimentaban, como mínimo de acuerdo a los lugares que fueron anunciados por los bailes. Para hacer esto, significa waggle duración de ejecución se estima para el primer baile que cada bailarín polen limitada realizada, que se comparó con la media correr duración durante los primeros bailes que realizan los trabajadores de ambos controles que bailaron durante las mismas horas de vídeo.
Además de forma natural forraje disponible, un pequeño número de trabajadores focales visitó un alimentador de sacarosa-solución que fue establecido para examinar la precisión de las carreras waggle realizadas por los trabajadores, ya que reclutaron a una fuente de alimento conocido en una ubicación fija. Precisión Danza se evaluó como la variabilidad en el ángulo y la duración de waggle corridas-sus componentes de dirección y distancia de un baile, respectivamente, para las danzas realizadas por los trabajadores de coordinación después de que regresaron desde el alimentador (medida como la desviación estándar de la media correr ángulo y la duración de cada baile, que luego se compararon entre tratamientos). Para garantizar que haya suficientes trabajadores de cada tratamiento visitado y bailado para el alimentador, introdujimos trabajadores adicionales en la colmena de observación de cada combinación de colonias / tratamiento en el ensayo 3. El número de etiquetas únicas era limitada, por lo que estos trabajadores adicionales fueron marcados con una pintura color que designa su origen (colonia y tratamiento de la fuente). Por lo tanto, la variabilidad en el ángulo de la danza se estimó para los trabajadores de pintura-marcados y etiquetados individualmente que visitaron el alimentador, mientras que todos los demás análisis del comportamiento de la abeja-baile (arriba) se calcula sólo para los trabajadores etiquetados. Debido a que la pintura marcado no podía distinguirse como individuos, bailes para el alimentador eran vistos como registros independientes. El alimentador se creó 188 m al norte de la colmena de observación y abastecido con solución de sacarosa 1,5 M o 2,0 M anís con aroma (dependiendo del interés forrajero). Abejas sin marcar fueron retirados del alimentador y enjaulados hasta que el alimentador se vació a limitar las visitas a sólo los trabajadores focales. Un observador en el alimentador transmitió las identidades de visitar trabajadores focales para el observador que estaba grabando la pista de baile y señalando sus danzas (que también eran distinguibles porque fueron generalmente similares). Los trabajadores que bailaban para el alimentador visitaron entre los días 19 y 29 de la prueba 3.
Enfoque estadístico
El número de trabajadores focales que hicieron o no participaron en una tarea se compararon entre los tratamientos con 2x2 (ensayo 1) o tablas de contingencia 2x3 (ensayos 2 y 3) el uso de pruebas de chi-cuadrado de independencia. Esperábamos que existirían diferencias en el peso del trabajador promedio y el rendimiento entre las colonias y entre los ensayos (que se realizaron durante los diferentes años y meses de verano). Sin embargo, nuestro enfoque no estaba en estas influencias temporales y genéticos en las métricas de los trabajadores, sino más bien de si los efectos del estrés polen a nivel de colonia se expresaron en forma consistente desempeño de los trabajadores en cada ensayo. Por lo tanto, se comparó el tratamiento significa separado para cada ensayo por t-test (prueba 1) o ANOVA (ensayos 2 y 3) y aplicó una corrección de Bonferroni para α = 0,05 para tener en cuenta para la realización de múltiples pruebas. Post-hoc de comparaciones por pares se realizaron utilizando el procedimiento de Tukey-Kramer, donde los efectos del tratamiento fueron significativos. La supervivencia de los trabajadores criadas en condiciones de tratamiento de polen limitado y polen abundantes se comparó con las estimaciones de Kaplan-Meier de supervivencia;comparaciones post-hoc se realizaron con ajustes Sidak para log-rank pruebas. T-pruebas, ANOVA y análisis de supervivencia se realizaron utilizando SAS versión 9.3 (SAS Institute, Inc., Cary, NC, EE.UU.); pruebas de contingencia se realizaron con una calculadora de acceso abierto estadística (physics.csbsju.edu/stats/contingency.ht ml) .El conjunto de datos está disponible como información de apoyo ( S1 Archivo ).
Resultados
Pesos adultos eran más bajos cuando las larvas se crían en colonias de polen limitado
En los tres ensayos, con un total de 1.808 trabajadores se introdujeron con éxito en las colmenas de observación (es decir, que estaban presentes durante al menos 24 horas después de la introducción de su colmena de observación: 638 trabajadores, criados en colonias cerradas polen limitada, 410 trabajadores criado en confinados colonias con abundante polen y 760 trabajadores criados en colonias no confinados con abundante polen).Los análisis para cada ensayo considerado aceptadas trabajadores focales solamente.
Trabajadores Focales criados en condiciones de limitación de polen habían reducido peso en comparación con los trabajadores que fueron criados en los controles confinados y no confinados ( fig. 1A ; prueba de 1: t 914 = 52.6, P <0 .0001="" 2:="" el="" ensayo="" f="" font="" nbsp="">2143 = 56.2, P < 0,0001; prueba de 3: F 2737 = 1336,5, P <0 font="" nbsp="">A través de los ensayos, los trabajadores que experimentaron la limitación del polen como larvas fueron 8-37% más ligero en la emergencia de adultos que a los trabajadores que fueron criados por compañeros de nido con acceso a abundante polen (controles). Las diferencias significativas entre los tratamientos de control de peso medio del surgimiento indicaron un efecto de ser criados por los trabajadores que estaban confinados a la colmena, incluso con abundantes suministros de polen ( Fig. 1A ).
La media (± SEM) A) peso fresco de los trabajadores de centros de coordinación en la emergencia de adultos, B) la longevidad de los trabajadores focales, C) la edad de inicio de la búsqueda de alimento para los trabajadores de centros de coordinación que se alimentaban, y D) el número de días observados de forrajeo de los trabajadores de centros de coordinación que se alimentaban. Trabajadores Focales se originaron a partir de colonias de origen que se dividieron en subunidades de colonias que tenían suministros limitados o abundantes de polen cuando los trabajadores focales fueron criados como larvas. Subunidades se circunscribían a una incubadora fría para evitar más forrajeo de polen (polen controles limitados o confinados) o habilitadas para alimentarse libremente (controles no confinados). Cuando el desarrollo se completa, los trabajadores focales fueron co-promovieron como adultos en una colonia del anfitrión no relacionado. El experimento se repitió en tres ensayos independientes que utilizan diferentes colonias de origen y de acogida. Las medias se hicieron comparaciones dentro de los ensayos donde los efectos del tratamiento fueron significativas después de la corrección de Bonferroni; diferencias significativas entre los tratamientos se indican mediante letras.
doi: 10.1371 / journal.pone.0121731.g001
En los tres ensayos, más trabajadores de las colonias de polen limitada se introdujeron sin éxito en su colmena de observación (es decir, que no se ve de nuevo después de 24 horas) que los trabajadores criados en las colonias de control confinados y no confinados (26% versus 16% y 12%, respectivamente ; todos los ensayos combinaron: χ 2 = 58,2, df = 2, P<0 font="" nbsp="">Esta diferencia fue consistente dentro de los ensayos (ensayo 1: χ 2 = 48,9, df = 1, P<0 .0001="" 2:="" el="" ensayo="" font="" nbsp="">2 = 26,6, df = 2, P <0 3:="" de="" font="" nbsp="" prueba="">2 = 35,8, df = 2 , P<0 font="">
Vida adulta era más corto cuando las larvas se crían en colonias de polen limitado
Longevidad de los adultos se redujo sustancialmente y constantemente cuando los trabajadores focales fueron criados como larvas en las colonias que estaban polen limitada en comparación con los trabajadores criados en colonias que tenían abundante polen ( Fig 1B. ; prueba de 1: t 919 = 7.1, P <0 .0001="" 2:="" el="" ensayo="" f="" font="" nbsp="">2143 = 6,7, P = 0,0016; prueba de 3: F 2737= 80.7, P <0 font="" nbsp="">La media de esperanza de vida se redujo como mínimo por 5 días y como máximo por 18 días para los trabajadores de polen limitados en comparación con los trabajadores de los grupos de control ( Fig. 1B ), lo que representa una disminución de la longevidad media de 21-56% entre los ensayos.
No hubo diferencias en la media de la longevidad trabajador entre los tratamientos de control en el ensayo 2 ( Fig. 1B ). En el ensayo 3, la longevidad media de los trabajadores fue de 3 días más corto para los trabajadores focales criados en colonias de control reducidos en comparación con los trabajadores criados en los controles no confinados ( fig. 1B ). Sin embargo, esta diferencia fue pequeña en comparación con la reducción de la longevidad que experimentan los trabajadores criados en colonias de polen y limitadas en el mismo juicio, vivieron 18 días menos en promedio en comparación con los trabajadores de los controles no confinados y 15 días menos que los trabajadores de controles confinados ( Fig . 1B ). La supervivencia de los trabajadores criados en colonias de polen limitada fue significativamente menor que la de los trabajadores de ambos tratamientos de control en cada ensayo ( Fig. 2 ; log-rank pruebas de función de supervivencia con ajustes Sidak; P <0 cada="" ensayo="" font="" para="">
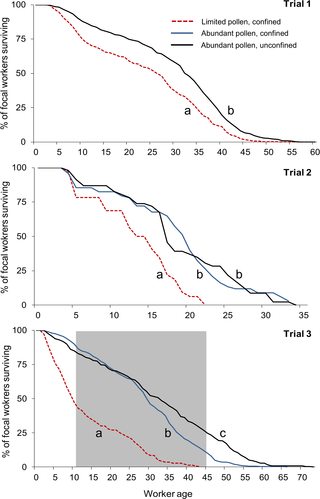 |
| Fig. 2 Supervivencia de los trabajadores adultos era más bajo cuando el acceso al polen se limitó durante el desarrollo larvario.
Los trabajadores fueron criados ya sea en colonias con polen limitada (y se limita a evitar una mayor búsqueda de alimento) o criados en colonias con abundante polen (ya sea confinado o permite que continúe en busca de alimento; controles). En cada ensayo, los trabajadores focales se introdujeron en una colmena de observación después de la emergencia de adultos; sólo aquellos trabajadores presentan 24 horas más tarde se incluyeron en el análisis de supervivencia. La zona gris de prueba de 3 indica que el período durante el cual se controló la contratación waggle-dance. Los datos brutos se representan, agrupados de diversas colonias dentro de un tratamiento por ensayo, en lugar de las estimaciones de Kaplan-Meier de la función de supervivencia (ver Métodos ). Las diferencias significativas en la supervivencia entre los tratamientos dentro de un ensayo se indican con letras diferentes.
doi: 10.1371 / journal.pone.0121731.g002
Actividad de forrajeo fue reducida cuando son criados en colonias de polen limitado
En los tres ensayos, la disponibilidad de polen durante la cría de larvas afectó significativamente el porcentaje de trabajadores que fueron observadas forrajeando como adultos (χ 2 = 65,6, df = 2, P <0 font="" nbsp="">Sólo el 62% de los trabajadores que fueron criados en colonias de polen limitada se observaron forrajeando en algún momento de su vida en comparación con el 81% y el 80% de los trabajadores criados en los controles suministrados en abundancia (tratamientos confinados y no confinados, respectivamente). Cuando se examina por separado, este resultado fue consistente en sólo dos de los tres ensayos después de una corrección de Bonferroni (ensayo 1: χ 2 = 40,8, df = 1, P <0 .0001="" 2:="" el="" ensayo="" font="" nbsp="">2 = 6,4, df = 2, P = 0,04 ; prueba de 3: χ 2 = 14,2, df = 2, P = 0,0008).
De los trabajadores de centros de coordinación que se alimentaban, las de cría en colonias de polen limitado tendían como adultos para comenzar a principios de forrajeo y consistentemente se alimentaron por menos días en comparación con sus homólogos de control. El inicio de la búsqueda de alimento fue acelerado por 2 días en promedio para los cazadores-recolectores de colonias de polen limitado en el ensayo 1 (t 657 = 3.0, P = 0,003; Fig. 1C ) y por 4-5 días en promedio en la prueba 3 (F 2,563 = 13.4, P <0 font="" nbsp="">Fig. 1C ). Forrajeo se inició a una edad similar en todos los tratamientos en el ensayo 2 (F 2109 = 0,7, P = 0,51; Fig. 1C ). Una vez forrajeo, los trabajadores criados en colonias de polen limitada se observaron en busca de alimentos en menos días que los trabajadores de los tratamientos de control: 1 día menos de media en el ensayo 1 (t 657 = 4.7, P <0 font="" nbsp="">la figura 1D. ), 2-3 días menos en el ensayo 2 (F2109 = 4,3, P = 0,016; Fig. 1D ), y 3 días menos en el ensayo 3 (F 2563 = 13.8, P <0 font="" nbsp="">Fig. 1D ).
En general, los cazadores-recolectores criados en colonias de polen limitado eran mucho más propensos a morir (es decir, desaparecen del registro) después de haber sido observado alimentándose únicamente para un solo día en comparación con los forrajeros criados en los controles suministrados en abundancia (30% de los cazadores-recolectores criados en polen limitado colonias desaparecieron después de un día contra 15% y 13% de los cazadores-recolectores de tratamientos de control confinados y no confinados, respectivamente; χ 2 = 51,6, df = 2, P <0 font="" nbsp="">Este efecto fue fuerte en dos de tres ensayos considerados aisladamente (ensayo 1: χ 2 = 28,9, df = 1, P <0 .0001="" 2:="" el="" ensayo="" font="" nbsp="">2 = 3,1, df = 2, P = 0,21; prueba de 3: χ 2 = 32,4, df = 2, P <0 font="">
Actividad Danza y precisión fueron reducidos cuando son criados en colonias de polen limitado
Un total de 397 danzas fueron realizadas por 116 trabajadores focales únicamente etiquetadas como visitaron una variedad de fuentes de alimentos durante el período de 32 días que la pista de baile se controló en el ensayo 3 (n = 9 bailarines criados en colonias cerradas polen limitado; n = 66 bailarines criados en colonias cerradas con abundante polen; n = 41 bailarines criados en colonias no confinados con abundante polen). De los trabajadores focales que fueron observados forrajeo, sólo el 9% de los trabajadores de cría en colonias de polen limitado, también se observaron baile comparado con el 24% y el 21% de los trabajadores criados en las colonias de control alimentados abundantemente confinados y no confinados (χ2 = 9,8, df = 2, P = 0,008; o 6% versus 19% y el 16% de los trabajadores que se introdujeron con éxito a la colmena, respectivamente: χ 2 = 14,2, df = 2, P = 0,001). Comenzamos grabar en vídeo la pista de baile tan pronto como observamos individuos focales bailando en los controles de la colmena frecuentes. Sin embargo, el porcentaje de trabajadores que bailó puede ser subestimado a través de tratamientos porque cualquier danzas realizadas por cazadores-recolectores que murieron antes de filmación comenzó no fueron registrados, y esta omisión pueden afectar desproporcionadamente a los trabajadores de menor duración que fueron criados en colonias de polen limitado (figs. 1B , 2 ). Si consideramos el grupo de bailarines potenciales a ser sólo los trabajadores que recolectaban durante el período en que la pista de baile fue grabado en vídeo, a continuación, la participación de baile seguía siendo bajo para los trabajadores criados en colonias de polen limitado, pero no de forma significativa (14% del polen trabajadores limitados frente a 28% y el 23% de los trabajadores de los controles confinados y no confinados; χ 2 = 4,6, df = 2, P = 0,10). Contrarrestando este argumento está el hecho de que los cazadores-recolectores criados en colonias de polen limitada se observaron baile con menos frecuencia y mucho más de ellos murieron después de ser observado forrajeando en un solo día (ver sección anterior), por lo que estos cazadores-recolectores tenían una menor probabilidad de contribuir a reclutamiento en el primer lugar.
Una vez que participan en el baile, el nivel de estrés de polen que los trabajadores experimentaron durante el desarrollo no afectó la cantidad de baile que lo hicieron, pero sí afectará a la precisión de sus danzas. Como trabajadores etiquetados forrajearon en las fuentes de alimentos desconocidos, la media del número total de días que cada bailarín bailó focal, el número total de danzas observados por bailarín durante esos días, el total de carreras waggle realizado, número de carreras waggle por la danza significa, y waggle significa -Ejecutar duración (un proxy para la distancia a las fuentes de alimentos anunciados) no fue diferente a través de tratamientos ( Fig. 3 ; F 2113 = 0,3, P = 0,72; F 2113 = 0,1, P = 0,88; F 2113= 0,6, P = 0,58; F 2113 = 1,1, P = 0,33; F 2,39 = 1,2, P = 0,32; respectivamente). Del mismo modo, la pintura marcado y etiquetado trabajadores de todos los tratamientos realizados números similares de carreras Waggle a su regreso desde el alimentador de sacarosa-solución ( Fig. 4 ; basado en el número de trabajadores pintados y etiquetados: 27 bailes realizados por al menos 6 diversos trabajadores criados en colonias de polen limitada, 63 danzas de al menos 18 trabajadores de diferentes colonias de control confinados, y 35 danzas de al menos 9 trabajadores de diferentes colonias de control no confinados; F 2122 = 2,2, P = 0,11). Sin embargo, los bailes de alimentación realizadas por los trabajadores de polen limitado transmiten información más variable sobre la dirección de la alimentación (es decir, los ángulos de carreras waggle en cada baile) que las danzas realizadas por los trabajadores de control (Fig. 4 ; F 2122 = 5,7, P = 0,002 ). La variabilidad en el componente de distancia de los bailes (es decir, las duraciones de pistas waggle en cada baile) fue similar entre tratamientos ( Fig. 4 ; F 2122 = 0,4, P = 0,88).
|
 |
| Fig. 3 comportamiento Waggle-dance de los trabajadores adultos no se vio afectada por el acceso al polen cuando los trabajadores focales eran larvas.
Siempre son medio por medidas bailarín de espectáculo de danza (± SEM) como individuos focales forrajearon en las fuentes de alimentos desconocidos en la prueba 3. Todos los trabajadores eran singularmente marcada e identificable individualmente.Actividad Waggle-dance se controló durante 1-2 h / día (como el clima y la búsqueda de alimento permitidos) desde el momento en que los trabajadores eran min. 12 días para máx. 45 días de edad (véase el recuadro gris en la Fig. 2 ). Las medias se calcula considerando sólo aquellos trabajadores que bailaban (es decir, no se incluyeron los valores de cero por falta de forraje o no bailar trabajadores focales; n = 9 trabajadores criados en colonias de polen limitado, confinado; n = 66 trabajadores criados en forma abundantemente, controles cerrados; n = 41 trabajadores criados en suministrados en abundancia, los controles no confinados). Duración waggle a ejecutar media (número de fotogramas a 30 cuadros por segundo, un proxy para la distancia a la fuente de alimentación anuncian) fue estimada para el primer baile realizado por cada trabajador polen limitado y en comparación con los medios para los primeros bailes que realizan los trabajadores de control durante la las mismas horas de cinta de vídeo (n = 9 trabajadores criados en subunidades de colonias de polen limitado, confinado; n = 20 trabajadores criados en controles abundantemente suministrados, confinados; n = 13 trabajadores criados en suministrados en abundancia, los controles no confinados).
doi: 10.1371 / journal.pone.0121731.g003

Fig 4. Los trabajadores criados en colonias con polen limitada realizó bailes waggle con mayor imprecisión direccional como adultos.
La variabilidad en la A) dirección y B) componentes distancia de danzas waggle se estimaron para los trabajadores que fueron criados ya sea en colonias con polen limitada (y confinadas para evitar una mayor búsqueda de alimento) o criados en colonias con abundante polen (ya sea confinado o permite que continúe forrajeo ; controles). Todos los bailes se realizaron para un alimentador de sacarosa-solución que se mantuvo a una ubicación fija de la colmena de observación en la prueba 3. bailes Feeder fueron realizados por trabajadores que tenían marcas de tratamiento específico (marcas de pintura y etiquetas), pero los individuos no eran siempre de forma única identificable, por lo que cada baile fue tratado como un registro independiente. Se calcularon las desviaciones estándar (DE) del ángulo y la duración de las carreras waggle para cada baile para comparar la precisión direccional entre tratamientos. Las diferencias entre tratamientos se indican con letras donde se encontraron efectos significativos de tratamiento.
doi: 10.1371 / journal.pone.0121731.g004
Discusión
El rendimiento de forraje y la contratación de trabajadores en la edad adulta se vio comprometida sustancialmente si se criaron como larvas en un entorno colonia polen limitado.En comparación con los compañeros de nido que fueron criados en condiciones de abundancia de polen, los trabajadores de polen estresadas eran más ligeras, que murieron antes, y menos de ellos se observaron forrajeo. Los que hicieron forraje inició forrajeo antes, se alimentaron por menos días, y tenían más probabilidades de morir después de un solo día de forrajeo. Además de ser menos probabilidades de forraje, los trabajadores criados en colonias de polen limitado también eran menos propensos a Danza de la abeja de los trabajadores de control y, si se bailaba, sus bailes eran menos precisas (aunque bailaban con un esfuerzo similar). Estos efectos sugieren un legado duradero para los trabajadores de estrés nutricional, que compromete seriamente la capacidad de búsqueda de alimento y el reclutamiento de los adultos, incluso cuando el estrés se limita sólo a la etapa larval. Es probable que estos déficits conductuales se agravarían si el estrés crónico persiste durante la edad adulta en el nivel de los trabajadores, y si una mayor proporción de trabajadores estaban desnutridos a nivel colonia. Estos efectos también pueden aumentar con el tiempo si los forrajeros estresados no pueden recolectar suficiente comida para las larvas de manera adecuada provisión, que luego pueden convertirse en adultos con bajo rendimiento a sí mismos. Dado el papel central que el juego de forrajeo y la contratación de la productividad en supervivencia de la colonia [ 78 - 80 ] y la utilidad de las abejas como polinizadores de cultivos [ 27 ], estos escenarios, con el apoyo de nuestros hallazgos, justifican las preocupaciones sobre el papel de la mala nutrición en declive colonia .
Forraje y reclutamiento son las tareas finales que las abejas realizan durante su vida, por lo que puede no ser sorprendente que los efectos acumulativos de estrés polen de desarrollo están fuertemente manifiestan en esta suite de punto final de comportamientos, y en formas matizadas. Efectos similares sobre la búsqueda de alimento y la contratación han sido inducida por otros factores de estrés ambientales en la vida de los trabajadores. La privación del sueño [81 ] y la baja temperatura de desarrollo [ 82 ] reducen la precisión de danzas waggle y, como en este estudio, estos efectos son a veces evidentes en sólo un elemento de la danza (es decir, la distancia frente a los componentes de baile) [ 82 ]. Baja temperatura de desarrollo [ 82] y la exposición a los pesticidas [ 83 ] también pueden reducir la probabilidad y el grado en que los trabajadores de la abeja de baile después de forrajeo. Si bien se sigue siendo posible que la probabilidad de baile para los trabajadores de polen estresadas fue subestimado en nuestro estudio, es probable que no lo era. No hay bailes por parte de trabajadores focales fueron observados antes del inicio de la grabación en vídeo de prueba de 3, a pesar de los controles repetidos cada día para determinar cuándo debe comenzar la grabación. Por otra parte, los trabajadores criados en colonias de polen limitado eran más propensos que los trabajadores de las colonias aprovisionados adecuadamente a desaparecer después de su primer día de búsqueda de alimento, lo que sugiere que pueden haber tenido dificultades para regresar a su colmena, posiblemente debido a una capacidad inferior para evadir la depredación, insuficiente vigor, pobre habilidad de orientación, u otra limitación física provocada por la desnutrición.Tales pérdidas para cosechadoras de forraje y la incapacidad para el hogar se encuentran también en los trabajadores que se ven afectados por otros factores de estrés ambientales (por ejemplo, plagas, patógenos, pesticidas, y los virus [ 84 - 88 ]). Mayor probabilidad de desaparecer después de un día de búsqueda de alimento significa que se espera proporcionalmente menos trabajadores estresados nutricionalmente a bailar, ya que nos encontramos. Por último, homing capacidad y eficiencia de forrajeo son altamente dependientes de los trabajadores de aprendizaje [ 89 ], que puede ser alterada en trabajadores desnutridas si tal estrés desarrollo interrumpe el aprendizaje de adultos y la memoria, al igual que en los vertebrados [ 8 , 90 - 93 ] y en moscas de la fruta [ 25 ]. Otros factores estresantes ambientales afectar el aprendizaje miel de abeja, incluyendo plagas y patógenos [ 94 , 95 ], la exposición a los pesticidas [ 83 , 96 ], y el estrés de temperatura [ 82 , 97 ], lo que sugiere la posibilidad de un efecto similar con el estrés nutricional.
Nuestros hallazgos confirman en una colonia natural de establecer el hallazgo consistente que el estrés polen de desarrollo reduce el peso y la longevidad de adultos cuando los trabajadores viven fuera de un contexto social (es decir, en jaulas) [ 71 , 73 - 75 ]. Los estudios sobre los efectos del estrés en los trabajadores de polen larvas criadas y que viven en las colonias no son comunes y tienden a no centrarse en el comportamiento de los trabajadores [ 69 ], con la excepción de la constatación de que el estrés polen larvas acelera la aparición de forrajeo [ 76] (que confirmado). El inicio temprano de la búsqueda de alimento para las personas estresadas desarrollo puede estar relacionado con relativamente rápido agotamiento de las reservas nutricionales, que precede el cambio de cubierta a las tareas al aire libre en las abejas de miel [ 98 ] y otros insectos sociales [ 99 , 100 ]. El acceso adecuado al polen puede aumentar la duración de la actividad de enfermería de interior [ 101 ], retrasando así el inicio de la búsqueda de alimento, pero la disponibilidad de polen fue equivalente para los co-promovido adultos en nuestro estudio, lo que significa que la transición del comportamiento de las tareas al aire libre se aceleró, en parte, por el mal estado nutricional de los trabajadores que estaban polen destacó como larvas (probablemente truncar la actividad de enfermería también).Además, se sugiere que los efectos conductuales de estrés nutricional de desarrollo no pueden ser totalmente rescatados por un acceso regular a alimentos más tarde en la vida.
Larvas de abejas están probablemente expuestos rutinariamente al estrés nutricional a corto plazo que se vivió por nuestros individuos focales, ya sea estacional o debido a las prácticas de gestión que limitan la disponibilidad de nutrientes. Esto es sugerido por la superposición entre el rango de peso para los trabajadores de polen-tónicas y átonas en este estudio (media de 71 a 113 mg al otro lado de los tratamientos y ensayos) y pesos reportados previamente para los trabajadores de emergencia de los adultos (81 a 140 mg; por [ 102 ]). Las larvas se someten a un aumento de peso 700 veces durante los 5-6 días que son amamantados [ 103 ], pero un solo día de mal tiempo reduce la actividad de enfermería por más de un medio, incluso cuando las colonias han almacenado polen [ 104 ]. Esta respuesta al mal tiempo probablemente explica por qué en nuestro estudio confinamiento solo (sin estrés polen) producido trabajadores que eran ligeramente más pequeño que las de cría en colonias no confinadas (pero con pocos efectos sobre el comportamiento). A largo plazo, el número de trabajadores en pequeñas colonias aumenta con episodios repetidos de mal tiempo [ 54 ] y los trabajadores más pesados se crían en momentos en que el polen está fácilmente disponible en una temporada [ 68 ], por lo que las diferencias entre los tratamientos en los pesos de emergencia probablemente reflejan los ajustes realizados en el aprovisionamiento de cría en colonias en respuesta a los cambios en ambos forrajeo de oportunidad y tiendas de polen (pero tenga en cuenta que el régimen no tuvo efecto sobre el comportamiento de los trabajadores de control alimentados en abundancia). Debido a la ventana en la que las larvas se alimentan es tan breve, del día a día los cambios en la atención de las enfermeras tienen el potencial de generar estrés nutricional para el desarrollo de los trabajadores y de los déficits correspondientes en función de adultos que hemos demostrado aquí. Vale la pena señalar que los trabajadores de polen limitado a menudo tenían un aspecto similar en tamaño a controlar a los trabajadores, por lo que sería difícil de determinar por inspección visual solamente que los trabajadores de colonias manejadas habían sido sometidos a estrés.
Un jefe de la preocupación por el impacto de la mala nutrición en las colonias de abejas de miel es la posibilidad de que actúa sinérgicamente con otros factores de estrés ambientales para socavar la función colonia. Cabe destacar que las larvas desnutridos son especialmente vulnerables a algunos de estos factores de estrés, incluyendo las plagas, patógenos y pesticidas. Los altos niveles de plaguicidas en el panal de cría durante el desarrollo larval reducen la longevidad de adultos [ 105 ], que probablemente se ve agravado por el estrés polen larvas debido a los pesticidas son más tóxicos para los deficientes en proteínas trabajadores [ 106 ]. Por otra parte, la exposición a plaguicidas de las células en el intestino medio, el sitio de la adsorción de nutrientes, aumenta la muerte celular en adultos y trabajadores de larvas [ 107 , 108 ], pero los efectos son especialmente pronunciada para las larvas. Susceptibilidad a los patógenos de cría de daño económico también se agrava cuando las larvas están desnutridos [ 72 , 109 , 110 ]. Por el contrario, las pupas parasitadas conVarroa destructor ácaros superar en parte los síntomas fisiológicos y de comportamiento de la infestación si se crían como larvas con acceso abundante al polen [ 76 ], pero los síntomas no se mitigan si una alimentación adecuada se entrega a los trabajadores sólo cuando son adultos [ 43 ]. Finalmente, los adultos infectados con el parásito intestinal Nosema exhiben muchos de los mismos síntomas que generamos con la limitación del polen larval solo (reducción de la longevidad, inicio temprano de forrajeo; [ 111 , 112 ]), así que estos dos factores estresantes pueden actuar sinérgicamente, a pesar de la exposición ocurre durante las diferentes etapas de la vida. Desafortunadamente, los síntomas de estrés polen larvas informaron aquí imitan los efectos negativos sobre los trabajadores de estos factores de estrés ambientales que también perjudican el acceso de los trabajadores a los nutrientes [ 107 , 108 ,113 - 115 ]. Juntos, múltiples factores estresantes pueden unirse en una tormenta perfecta de condiciones que hacen que sea difícil para los trabajadores adquieren el alimento que necesitan para funcionar recolectores y bailarines como eficientes. Esta posibilidad, que es sugerido por nuestros resultados, justifica la preocupación por el efecto de la mala nutrición en la salud y la productividad de las colonias de abejas de miel y la calidad de los servicios de polinización que pueden ofrecer.
Agradecimientos
Estamos muy agradecidos a Amanda Gardner, Rachel Reed, Catherine Oleskewicz, Anita Yau, y Amina Ziad para obtener ayuda con el etiquetado de las abejas, la recogida de datos de las colmenas de observación y análisis de vídeo.
Contribuciones de los autores
Concebido y diseñado los experimentos: HRM. Realizado los experimentos: HRM SNP.Analizados los datos: HRM SNP. Aportados reactivos / materiales / herramientas de análisis: HRM. Escribió el documento: HRM SNP.
Referencias
|


No hay comentarios:
Publicar un comentario