Contaminación acústica altera servicios ecológicos: una mayor interrupción de la polinización y dispersión de semillas.
1. Clinton D. Francisco 1 , * ,
2. Nathan J. Kleist 2 ,
3. Catherine P. Ortega 3 y
4. Alejandro Cruz 2
+Afiliaciones de los autores
1. ↵ * Autor para la correspondencia ( clinton.francis @ nescent.org ).
Abstracto
La contaminación acústica es una fuerza nueva, medioambiental generalizada que recientemente se ha demostrado que alteran el comportamiento y distribución de las aves y otros vertebrados, pero si el ruido tiene acumulados, a nivel comunitario mediante el cambio de consecuencias importantes servicios ecológicos es desconocida. En este documento, hemos examinado los efectos de la contaminación acústica sobre polinización y la dispersión de semillas y el establecimiento de plántulas dentro de un sistema de estudio que aisló los efectos del ruido de los estímulos de confusión comunes a los humanos alterados paisajes.Utilizando observaciones, encuestas vegetación y la transferencia de polen y los experimentos de remoción de semillas, se encontró que los efectos de la contaminación acústica pueden repercutir en las comunidades mediante la interrupción o la mejora de los servicios ecológicos. En concreto, la contaminación acústica indirectamente aumentado polinización flor artificial por los colibríes, pero alteró la comunidad de animales que se alimentan y se dispersan sobre Pinus edulis semillas, lo que podría explicar reducido P.edulis reclutamiento de plántulas en áreas ruidosas. Aunque está claro que algunos servicios ecológicos, tales como la polinización, pueden beneficiarse indirectamente debido al ruido, a la disminución en el reclutamiento de plántulas para la clave que dominan especies como P. edulis puede tener graves efectos a largo plazo sobre la estructura de los ecosistemas y la diversidad. Debido a la magnitud de la contaminación acústica es cada vez mayor, este estudio pone de relieve que los investigadores deben evaluar las consecuencias ecológicas de ruido, junto con otros cambios antropogénicos del medio ambiente que están remodelando el hombre paisajes alterados por todo el mundo.
1. Introducción
Las actividades humanas han alterado más del 75 por ciento de la superficie terrestre de la Tierra [ 1 , 2 ]. Concomitante con estos cambios en la superficie es un aumento generalizado en el ruido antropogénico, o la contaminación acústica causada por la expansión de redes dendríticas transporte, centros urbanos y las actividades industriales [ 3 ]. La extensión geográfica de la exposición al ruido varía según la región y la escala, pero se estima que una quinta parte de la superficie terrestre de los Estados Unidos se ve afectado por el ruido del tráfico directamente [ 4 ] y más del 80 por ciento de unos paisajes rurales están expuestos a niveles de ruido mayores debido a las actividades de extracción de energía [ 5 ]. A pesar de la escala potencialmente importante de la exposición al ruido en el mundo, muy poco se sabe acerca de cómo estas nuevas condiciones acústicas ecológicamente afectar a las poblaciones naturales y las comunidades.
Estamos empezando a comprender los efectos de la exposición al ruido creciente en los comportamientos de los individuos y las distribuciones de las especies [ 6 - 10 ], y varios estudios recientes contorno potencial y algunos efectos conocidos de ruido [ 3 , 11 - 13 ]. A pesar de esta reciente atención dada a los efectos del ruido, todavía tenemos un conocimiento limitado de cómo estos impactos a escala para los procesos comunitarios y de los ecosistemas nivel. Algunos estudios han demostrado que los depredadores evitar zonas ruidosas [ 7 , 14 - 16 ], presumiblemente debido a que deteriora la capacidad de ruido depredadores para localizar a sus presas. Estos estudios nos proporcionan ideas sobre cómo el ruido puede afectar directamente a interacciones depredador-presa, pero no proporcionan información sobre si el ruido puede tener consecuencias acumulativas, indirectas para otras interacciones y organismos que no se ven afectadas por el ruido directamente.
Nuestro objetivo fue investigar si la contaminación acústica pueden repercutir en las comunidades ecológicas, afectando especies que proporcionan servicios ecológicos funcionales únicas. Nos hemos centrado nuestros esfuerzos en los servicios ecológicos proporcionados principalmente por las aves, ya que se considera que son especialmente sensibles a la contaminación acústica, debido a su dependencia de la comunicación acústica [ 11 ]. Sin embargo, como no todas las especies responden de manera uniforme a la exposición al ruido [ 6 , 7 , 10 , 17 ], podemos evaluar cómo las diferentes respuestas funcionalmente por organismos únicos impacto indirecto de otras especies y desencadenar otros cambios en la estructura de la comunidad. Estudiamos los servicios ecológicos proporcionados por Arquíloco alexandri (negro-chinned hummingbird) y Aphelocoma californica (oeste de fregar-jay), que sirven como vínculos móviles para la polinización y Pinus edulis (piñón) Servicios de dispersión de semillas, respectivamente [ 18 - 20 ]. Debido a A. Alexandripreferentemente nidos en ambientes ruidosos y A. californica evita las zonas ruidosas [ 5 , 7 ], se propuso que sus dependientes ruido distribuciones podría dar lugar a una mayor tasa de polinización de plantas polinizadas por colibríes y perturbar P. edulis servicios de dispersión de semillas en zonas ruidosas y potencialmente afectan el reclutamiento de plántulas (figura 1 a ).
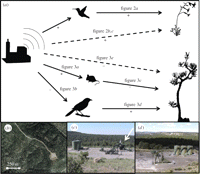
Ver una versión más grande:
· Descargar como diapositiva de PowerPoint
Figura 1.
( a ) Camino por las que el ruido altera los servicios de polinización y dispersión de semillas. Las flechas continuas y discontinuas denotan las interacciones directas e indirectas, respectivamente. Señales de referencia para efectuar la dirección, y el apoyo para cada efecto se indica mediante el número de figura. Véase el texto principal de los resultados y las citas que apoyan la dependencia de I. aggregata en A. alexandri (flecha etiquetada figura 2 una ) y para la calidad funcional de Peromyscus ratones y A. californicacomo P. edulis dispersores de semillas (flechas etiquetadas figura 3 c , d ). ( b ) los pozos activos de gas situadas en el extremo de las vías de acceso sirve como ( c ) los sitios de tratamiento ruidosos debido a la presencia de gas de generación de ruido y compresores (flecha blanca) o ( d ) los sitios de control tranquilas.
Para probar estas predicciones, se utilizó un sistema único de estudio que aísla la influencia de la exposición al ruido de muchos factores de confusión comunes a las zonas ruidosas, tales como heterogeneidad vegetación, los efectos de borde y la presencia de los seres humanos y los vehículos en movimiento (ver más abajo). Utilizamos observaciones, encuestas vegetación y la transferencia de polen y los experimentos de remoción de semillas en pares de sitios de tratamiento y control para determinar cómo difieren las interacciones ecológicas en áreas ruidosas y tranquilidad y si el ruido afecta indirectamente a las plantas que dependen funcionalmente únicos enlaces móviles de aves.
2. Material y métodos
Nuestro estudio se llevó a cabo en el Área de Manejo de Hábitat Rattlesnake Canyon (RCHMA), ubicada en el noroeste de Nuevo México.Estudio zona detalles y el sitio se puede encontrar en otro lugar [ 5 - 7 ]. En pocas palabras, RCHMA está dominado por el arbolado formado por P. edulisy Juniperus osteosperma (enebro) y tiene una alta densidad de pozos de gas natural ( figura 1 b ). Muchos pozos están acoplados con los compresores que funcionan de forma continua y generar ruido a amplitudes altas (mayores de 95 dB (A) a una distancia de 1 m), y, como la mayoría de ruido antropogénico, el ruido del compresor tiene una energía considerable a bajas frecuencias y disminuye hacia mayor frecuencias (material complementario electrónico, figura S1) [ 5 - 7 ]. Además, la actividad humana en los pozos y las características principales de la vegetación en los bosques que rodean los pozos no difieren entre los pozos con (sitios ruidosos tratamiento) y sin compresores generadores de ruido (sitios tranquilos de control, figura 1 c, d ) [ 7 ], que proporcionan una oportunidad para evaluar el efecto indirecto de ruido en el apoyo a los servicios ecológicos en la ausencia de muchos estímulos de confusión comunes a la mayoría de los humanos alterados paisajes.
(A) La polinización experimento
Para determinar si los colibríes flores polinizadas indirectamente se benefician de ruido, se utilizó un experimento de campo para el control de la densidad y la distribución espacial de los recursos de néctar colibrí con parches de flores artificiales que imitaban una planta de auto-incompatible, el colibrí de polinización comunes a nuestra área de estudio : Ipomopsis aggregata (material complementario electrónico, cifras S2 y S3 a ). En mayo de 2010, se establecieron siete pares de sitios de tratamiento y de control dentro de RCHMA para los experimentos de polinización. Los sitios fueron emparejados geográficamente para minimizar las posibles diferencias en características de la vegetación dentro de cada par, sin embargo, para asegurar que los niveles de ruido de fondo fueron significativamente diferentes entre pares de sitios, los sitios fueron mayores que o igual a 500 m de distancia y resultó en condiciones relativamente tranquilas en los sitios de control. La distancia resultante entre pares de tratamiento y control fue de 767 m (± 57 sem, mínimo = 520 m, máximo = 954 m).
Parches de flores artificiales se crearon 125 m, ya sea la cabeza del pozo o el compresor en el control y centros de tratamiento, respectivamente (material complementario electrónico, figura S2 a ). La dirección del primer parche con relación a la cabeza del pozo o el compresor se determinó al azar y el segundo parche se estableció 40 m de la primera y también a 125 m de la cabeza de pozo o un compresor. Antes del experimento, en cada parche, que mide la amplitud de ruido de fondo como decibelios ponderados (dB (A)) 1 min para confirmar que los niveles de ruido fueron significativamente mayores en los parches relativos del tratamiento para controlar los parches. En todos los casos, las mediciones en el tratamiento emparejado y sitios de control se realizaron en el mismo día y en aproximadamente el mismo tiempo. Se midió la amplitud como el nivel de ruido continuo equivalente ( L EQ , rápido tiempo de respuesta) con metros Casella convertibles de presión de sonido del dosímetro / sonido (modelo CEL 320 y el convertidor de CEL 1002). Se utilizó 95 mm parabrisas acústico, y no tomamos medidas cuando las condiciones del viento se clasificaron tres o más en la escala de Beaufort del viento (aprox. 13-18 km h -1 ), o cuando suena distinto compresor de ruido (es decir, vocalizaciones de aves y ruido de las aeronaves) podría sesgar las mediciones.
Flores artificiales se utilizan frecuentemente en estudios de polinización [ 21, 22 ] y los utilizados en nuestro experimento se construyeron a partir de 0,6 ml tubos de microcentrífuga. Este tamaño de tubo de microcentrífuga había sido utilizado previamente en experimentos de polinización con A.alexandri [ 23 ]. Para imitar la apariencia de I. aggregata , nos envuelve cada tubo de microcentrífuga con cinta roja eléctrica (material complementario electrónico, S3 figura). Además, se adjuntan tres pequeñas piezas de hilo de color amarillo para proporcionar un sustrato para el marcado de flores con transferencia de tinte fluorescente y posterior y la deposición en las flores por otros polinizadores. Cada planta artificial consistía en tres flores unidos a una barra de metal 53 cm de largo con cinta aislante verde (material complementario electrónico, figura S3 b ). Los parches de plantas fueron dispuestas en 3 m 2 zona con cuatro plantas que marcan cada esquina y uno en el centro (material complementario electrónico, figura S2 a).
Parches de plantas se establecieron simultáneamente o uno inmediatamente después del otro (menor o igual a 30 min) en pares de sitios. Debido I. aggregata néctar es de 20-25% de sacarosa [ 24 , 25 ], nos llena cada flor con una recompensa de 0,4 ml de solución de sacarosa al 25 por ciento con pipetas calibradas y goteros de plástico, volviendo todos los días a la misma hora aproximadamente para volver a llenar la flor con la sacarosa recompensa para que los polinizadores aprendido a utilizar las flores como recurso forrajero. Sólo rara vez nos encontramos con una sola flor artificial completamente agotado de la recompensa entre las visitas para reponer la recompensa, pero nunca las tres flores de la misma planta.
Hemos realizado observaciones para determinar las tasas de visitas de polinizadores en 11 (79%) de los 14 pares de parches de tratamiento y control. Debido a que la creación de nuestros parches llevó varios días, antes de que nuestras observaciones, cuatro pares de parches fueron rellenados durante 4 días, dos parches fueron rellenados por 3 días y cinco parches fueron rellenados por 2 días y todas las revisiones observadas se había establecido durante más de 38 h antes de la observación. A continuación, realizó observaciones en parches en pares de sitios de control y de tratamiento simultáneamente o uno inmediatamente después del otro.Vimos flores en parches de coordinación de 15 minutos y marcó el número de visitas a cada planta a una distancia de 5 m, usando binoculares cuando sea necesario para identificar artrópodos que visitan las flores. Todos los polinizadores no colibrí se separaron en sus órdenes (Diptera Hymenoptera y Lepidoptera) y se utilizó Poisson generalizados lineales mixtos modelos (GLMM) dentro del paquete lme4 en R [ 26 ] para examinar si las visitas de conexión por A. alexandri o otros polinizadores difieren entre el tratamiento y sitios de control. Los sitios individuales y sitios geográficamente pareadas fueron tratados como efectos aleatorios.
A raíz de las observaciones de actividad, el 28 de mayo de 2010, volvimos a todos los programas entre las 07.00 y las 12.00 para volver a llenar todas las flores artificiales con la recompensa sacarosa y singularmente marcado por una planta de parche con el polvo fluorescente amarillo o rojo (Day-Glo Color Corporation, Cleveland, Ohio, EE.UU.) de tal manera que las plantas en el mismo lugar pero en diferentes parches recibido un polvo de color único. El uso de polvo fluorescente como un proxy para la transferencia de polen es una técnica ampliamente utilizada en los estudios de polinización porque la transferencia del polvo se correlaciona fuertemente con la transferencia de polen [ 27 , 28 ]. Cada parche se permitió 24 h de exposición para visitas de polinizadores antes se recogieron cada planta para su posterior examen para la transferencia de polvo en el laboratorio.
En el laboratorio, se utilizó una lámpara ultravioleta bajo condiciones de oscuridad para registrar la presencia o ausencia de polvo en cada inflorescencia, observando si el polvo de la planta fue marcado en el mismo parche o el parche situado a 40 m. A continuación, utiliza Poisson GLMMs a examinar dentro de la parcela y entre-patch transferencia de polen con el número de flores individuales por parche con polen transferido como variables de respuesta. Tratamos a cada lugar geográfico y el tratamiento emparejado y sitio de control como efectos aleatorios en todos los modelos.
(B) Pinus edulis semillas eliminación experimento
Hemos llevado a cabo P. edulis remoción de semillas en todo RCHMA experimentos para determinar si y cómo las tasas de remoción de semillas y la comunidad de depredadores y dispersores de semillas responder a la exposición al ruido. Pinus edulis árboles dentro de una región típicamente sincronizar la producción de cultivos a gran cono cada 5-7 años [ 20 ]. Como conos gradualmente secos y abiertos en septiembre, no se recogen las semillas de los conos de córvidos en el otoño dosel hasta el suelo, donde los roedores, córvidos y otras especies de aves y consumir semillas de cosecha durante varios meses [ 20 ]. Las tasas de seguimiento de mudanzas en otoño de semillas del suelo puede ser problemático, ya que las semillas siguen cayendo de los árboles, por lo tanto, llevamos a cabo nuestros experimentos en junio y julio, cuando ningún otro P. edulis semillas estaban disponibles, de forma similar a otros estudios que han examinadoP. edulis remoción de semillas y su dispersión durante los meses de verano [29 ].
Utilizamos seis pares de tratamiento y control de sitios que fueron unidas geográficamente. Sitios cumplido esos mismos criterios descritos para el experimento de polinización. La distancia media entre pares de tratamiento y control fue de 821 m (± 51 sem, mínimo = 642 m, máximo = 1029 m). En cada sitio, se establecieron estaciones de siembra en 10 lugares a menos de 150 m de cada bien o compresor (material complementario electrónico, figura S2 b ). Ubicaciones fueron seleccionados al azar, siempre que la distancia entre cada estación fue mayor o igual a 40 m, y cada estación se encuentra en el suelo bajo una madurez reproductiva P. edulis árbol.
La remoción de semillas experimentos duró 72 horas con visitas a cada estación cada 24 h para cuantificar la tasa diaria de remoción de semillas. Al comienzo de cada período de 24 horas, que simula la caída natural de la semilla por la dispersión de 20 P. edulis semillas en el suelo en un 0,125 m 2zona. Después de regresar 24 horas después de documentar el número de semillas removidas, determinar si existían pruebas para in situ depredación de semillas por buscar cuidadosamente el área inmediata (aprox. 2 m 2 ) para el recién inaugurado P. edulis semillas (generalmente visible como una colección agrupada de fragmentos de cubierta de la semilla-de varias semillas) y de nuevo a la dispersión de las semillas 20 en la estación.Evidencia de depredación de semillas en una estación se define como si abiertos recientemente (y vacío) cubiertas de las semillas se detectaron en ninguna de las tres visitas utilizados para cuantificar las semillas tasa de remoción. Todas las semillas fueron recolectadas a nivel local dentro RCHMA y fueron manejados con guantes de látex para que el olor humano no se ha transferido a las semillas. Durante una de las cuatro visitas a cada estación, que mide la amplitud de ruido de fondo siguiendo los métodos descritos anteriormente para el experimento de polinización.
Para documentar la identidad de los animales de la eliminación de las semillas, que combina cada estación con un movimiento-activado cámara digital (Wildview Xtreme II). Las cámaras fueron montadas en un tronco o una rama de un árbol adyacente dentro de 1-3 m de la estación de semilla de una visión clara, sin embargo, colocarse en un lugar relativamente discreto para no llamar la atención adicional a la estación. Cámaras permaneció en cada estación durante todo el periodo de 72 horas y se documentó la remoción de semillas, tanto diurnos como nocturnos. Una detección positiva de una especie eliminación de semillas se registra sólo cuando un individuo se documentó retirar o consumir semillas.
El número de semillas removidas por 24 h del periodo se utilizó para calcular una proporción diaria promedio de semillas eliminado, lo que nos arcoseno raíz cuadrada transformado para cumplir los supuestos de normalidad y homogeneidad de la varianza. Se utilizó lineales mixtos modelos (LMMS) para examinar si la proporción de semillas removidas difería entre el tratamiento y las estaciones de control o de siembra debido a la presencia o ausencia de las especies individuales. Se utilizó GLMMs binomiales para determinar la presencia de especies individuales explicado in situdepredación de semillas, evidenciada por la presencia de reciente apertura,P. edulis testa. Para los modelos en los que hemos examinado la influencia de las distintas especies en la remoción de semillas o la depredación, empezamos con los modelos que contienen todas las especies documentadas como variables predictoras y procedió a retirar cada una de las variables no significativas a la vez sobre la base de los más altos p -valor hasta sólo efectos significativos se mantuvo. Se utilizó Poisson y binomial GLMMs examinar si la riqueza de especies de la comunidad y la eliminación de detecciones de semillas de especies individuales difieren en las estaciones de tratamiento de semillas y control, respectivamente. Para todos los modelos, tratamos cada sitio y tratamiento geográficamente vinculados y centros de control como efectos aleatorios. Algunos modelos de evaluación de las detecciones de las especies individuales de tratamiento y los sitios de control no convergen, por lo tanto, para estos casos, se utilizó χ 2 : pruebas para determinar si existe una diferencia entre el número total de detecciones en sitios de control y de tratamiento.
(C) encuestas reclutamiento de plántulas
En 2007, completamos 129 estudios de vegetación al azar en 25 parcelas de vegetación m de diámetro (aprox. 490 m 2 ) ubicados en nueve tratamientos y ocho sitios de control, algunos de los cuales no eran los mismos lugares utilizados en los experimentos de remoción de semillas, que incluyeron sólo seis pares de sitios (12 en total). Diez centros de tratamiento fueron encuestados en 2007, pero se excluyeron parcelas de vegetación de un lugar a partir de este análisis, ya que el compresor se instaló en 2006, con lo que confundir las condiciones acústicas en los que muchas plántulas pueden haber sido establecidas (ver más abajo).Compresores en todos los centros de tratamiento más habían estado en vigor durante al menos 6 años, pero a lo largo de 10 años para varios sitios.
Debido a que nuestro trabajo de campo en los años anteriores había documentado una evitación de ruido por A. californica [ 7 ], en el año 2007, contamos con todo P. edulis plántulas por parcela vegetación. Nos cuenta restringidas de plántulas a aquellos. Menor o igual a 20 cm para asegurarse de que se había dispersado y estableció hace relativamente poco y bajo las mismas condiciones acústicas que estaban presentes en 2007 Asumimos plántulas de menos de o igual a 20 cm se había dispersado y establecido en los últimos 6 años debido a 1 año de edad, P. edulis plántulas se midió a tener una altura promedio de 5,3 cm [ 30 ] y porque las estrechamente relacionadas con Pinus cembroides alcanza una altura de 1 m alrededor de los 5 años de edad [ 31 ]. Por lo tanto, nuestros supuestos deben ser considerados conservadores. Analizamos el reclutamiento de plántulas con el número de plantas por parcela como la variable de respuesta utilizando GLMMs Poisson. Las variables predictoras incluidas ubicación de la parcela ya sea en un tratamiento o de control, sino también las características a nivel del lote que pueden influir en el establecimiento de plántulas y de contratación, tales como el número de arbustos, P. edulis y J. osteospermaárboles, el monto de la cobertura del dosel, la profundidad de la hojarasca y el porcentaje de cobertura del suelo clasificado como material vivo, la materia muerta o suelo desnudo. Identidad del sitio fue tratado como un efecto aleatorio. Seguimos el mismo procedimiento de selección descrito anteriormente para la remoción de semillas y depredación de semillas. Los datos para el reclutamiento de plántulas, además de los datos de la remoción de las semillas y experimentos de polinización se han depositado en Dryad ( www.datadryad.org ; doi: 10.5061/dryad.6d2ps7s7).
3. Resultados
(A) La polinización
Los valores de ruido de amplitud fueron significativamente mayores (aprox. 12 dB (A)) en los parches relativos del tratamiento para controlar los parches (LMM: χ 2 1 = 25,550, p <0,001, el material suplementario electrónico, figura S2 c ) y experimentado similares a los aproximadamente 500 metros de las autopistas [ 32 , 33 ]. Observaciones focales en un subconjunto de los parches reveló que varios taxones visitado flores artificiales se suministran con una recompensa de néctar (material complementario electrónico, tabla S1), sin embargo, sólo A. Alexandri visitas difieren entre el tratamiento y sitios de control. Archilochus Alexandri visitas eran cinco veces más comunes en los parches de tratamiento que los parches de control (Poisson GLMM: χ 2 1 = 6,859, p = 0,009, figura 2 un ).

Ver una versión más grande:
· Descargar como diapositiva de PowerPoint
Figura 2.
La evidencia de experimento de polinización. ( a )Archilochus alexandrivisitado flores artificiales en los parches de tratamiento ruidosas más silenciosas que los parches de control in situ. Los valores mostrados reflejan la cantidad de visitas por sitio (14 sitios en total con dos muestras de cada sitio, cinco plantas por parcela y tres flores por planta). ( b , c ) polinización de las flores individuales fue más alta en los sitios de tratamiento en relación a controlar tanto los sitios ( b ) dentro de y ( c ) entre los parches.Los valores mostrados reflejan la suma de las flores polinizadas en ambas muestras de cada sitio. ( a - c ) Líneas de vincular sitios geográficamente vinculados.
Consistente con más A. alexandri visitas a las plantas en las zonas ruidosas, dentro del parche de transferencia de polen se produjo el 5 por ciento de las flores de control del sitio, pero el 18 por ciento de las flores del sitio de tratamiento (Poisson GLMM: χ 2 1 = 15,518, p <0,001, Figura 2 b ) y entre-patch transferencia de polen se produjo el 1 por ciento de las flores de control del sitio y el 5 por ciento de las flores del sitio de tratamiento (Poisson GLMM: χ 2 1 = 6,120, p = 0,013, figura 2 c ). Los análisis mediante la presencia o ausencia de polen transferido al nivel de parches reveló el mismo patrón (dentro del parche GLMM binomio: χ 2 1 = 8,800, p = 0,003; entre-patch GLMM binomio: χ 2 1 = 5,608, p = 0,018) .
(B) Pinus edulis remoción de semillas
Los valores de ruido de amplitud fueron consistentemente más altas (aprox. 14 dB (A)) en las estaciones de tratamiento de semillas con relación a controlar las estaciones de siembra (LMM: χ 2 1 = 19,084, p <0,001, el material suplementario electrónico, figura S2 d ), sin embargo, la semilla ni velocidad de eliminación (LMM: χ 2 1 = 2,209, p = 0,137), ni riqueza documentado especies por estación semilla diferente entre sitios con y sin ruido (Poisson GLMM: χ 2 1 = 0,461, p = 0,497).
La mayoría de los animales detectados con cámaras activadas por movimiento, la eliminación de semillas de las estaciones fueron identificados fácilmente a las especies, sin embargo, para los dos grupos, Peromyscusratones y Sylvilagus conejos, no siempre fueron capaces de identificar a los individuos de las especies, por lo que fueron asignados a su géneros respectivos. En total, documentamos 11 semillas taxones de depilar, nueve de los cuales fueron considerados depredadores de semillas (material complementario electrónico, tabla S2). Cámaras no pudo detectar la identidad de los animales que eliminaron las semillas aproximadamente a una estación por el sitio, principalmente debido a una falla de pila. Sin embargo, no hubo diferencia en el número de fallos de la cámara entre el tratamiento y los sitios de control que sugieren nuestros detecciones estaban sesgadas hacia el tipo de sitio uno sobre el otro (GLMM binomial: χ2 1 = 0,240, p = 0,624), por lo tanto, cualquier pariente diferencias en las detecciones entre el tratamiento y los centros de control debe reflejar diferencias reales entre las zonas ruidosas y tranquilo.
De los nueve depredadores de semillas documentados quitar las semillas, sólo uno, Pipilo maculatus , se detectó con mayor frecuencia en los sitios de control en relación con los centros de tratamiento (binomio GLMM: χ 2 1 = 4,133, p = 0,042), un patrón consistente con hallazgos anteriores de que P.maculatus evita el ruido en su colocación nido [ 7 ]. También documentamos la remoción de semillas por Peromyscus ratones y A. californica , que se considera principalmente depredadores de semillas y dispersores importantes de semillas, respectivamente [ 20 ]. Los ratones se detectó en el 63 por ciento de las estaciones de tratamiento de semillas y sólo el 45 por ciento de estaciones de control (binomial GLMM: χ 2 1 = 4,023, p = 0,045; figura 3 una ). Por el contrario, A. californica se detectó la eliminación de semillas exclusivamente en las estaciones de control ( χ 2 1 = 5,486, p = 0,019; figura 3 b ). Peromyscus ratones y A. californica fueron también los únicos taxa con fuertes efectos sobre la remoción de semillas y, junto conlos requisitos minimos Tamias , fueron los taxones con una fuerte influencia sobre los patrones de depredación de semillas en la estación de siembra (es decir, presencia de cubiertas de las semillas abiertas). Las tasas de remoción de semillas fue de aproximadamente 30 por ciento mayor en las estaciones donde Peromyscus ratones o A. californica fueron documentados quitar las semillas en comparación con las estaciones en las que no fueron detectados (LMM: χ 2 2 = 35,775, p <0,001; figura 3 c, d ). Depredación de semillas fue afectada positivamente por la presencia de Peromyscus ratones ( β ratón = 0,841 ± 0,412 se) y T. minimos ( β chipmunk = 1,199 ± 0,544 SE) ,ambos depredadores de semillas típicamente considerados [ 20 ], pero negativamente afectada por la presencia de A. calilfornia ( β -scrub jay = -2,031 ± 1,005 se; GLMM binomio: χ 2 3 = 13,748, p = 0,003). De hecho, la mayoría de las estaciones donde Peromyscus ratones (74%) y T. requisitos minimos (81%) se detectaron también tenían evidencia de depredación de semillas, pero sólo el 33 por ciento de las estaciones en las que A. Californiase detectó hubo signos de depredación de semillas.

Ver una versión más grande:
· Descargar como diapositiva de PowerPoint
Figura 3.
( a ) Peromyscus ratones se detectaron con más frecuencia en las estaciones de tratamiento de semillas con relación a las estaciones de control. (b ) A. californica eliminado semillas exclusivamente en los sitios de control. Los valores mostrados en ( a ) y ( b ) reflejan la proporción de estaciones de semillas por sitio donde los ratones o los arrendajos se han detectado. ( c , d ) las tasas de remoción de semillas (por 24 h) fueron mayores cuando ( c ) Peromyscusratones o ( d ) A. californica se detectaron en una estación de semilla. ( e ) Pinus edulis reclutamiento de plántulas fue significativamente mayor en los sitios de control en relación con los centros de tratamiento. Los diagramas de caja indican el valor de la mediana (línea de color negro sólido), 25 y percentiles 75 (recuadro), y los bigotes denotar el rango intercuartil 1,5. Los valores atípicos se denotan por los círculos abiertos.
(C) Pinus edulis reclutamiento de plántulas
De acuerdo con la diferencia en la eliminación de los animales semillas en las zonas ruidosas y tranquilo, P. edulis plántulas tenían cuatro veces más abundantes en los sitios de control en relación con los centros de tratamiento ( β tratamiento = -1,543 ± 0,240 se; figura 3 e ), pero el número de J. osteosperma árboles ( β Juniper = 0,036 ± 0,016 se) y la proporción de cobertura muerta del suelo orgánico ( β Dead = 0,023 ± 0,008 se) tuvo efectos pequeños y positivos sobre la abundancia de plántulas (Poisson GLMM: χ 2 3 = 38,583, p <0,001) . Sin embargo, ninguna de estas variables, ni el número de P. edulis árboles, difieren entre el tratamiento y los sitios de control (enebro LMM: χ 2 1 = 0,726, p = 0,394; muerto cobertura del suelo LMM: χ 2 1 = 0, p = 1,0; P. edulis LMM: χ 2 1 = 2,560, p = 0,110), lo que sugiere que otras características del hábitat puede ser excluido como explicaciones alternativas para P. edulis reclutamiento de plántulas en el tratamiento y sitios de control.
4. Discusión
Los niveles elevados de ruido afectado las tasas de polinización por colibríes y P. edulis dispersión de semillas y el reclutamiento de plántulas, pero la dirección de cada efecto era diferente. La exposición al ruido tiene un efecto positivo indirecto sobre la polinización por colibríes, pero un efecto negativo indirecto sobre P. edulis establecimiento de las plántulas mediante la alteración de la composición de los animales cazando sobre o dispersión de semillas. Estos resultados amplían nuestro conocimiento de las consecuencias de la exposición al ruido, que se ha centrado principalmente en las respuestas vocales a ruido [ 8 , 34 , 35 ], tanto en la distribución de especies y el éxito reproductivo [ 7 , 10 , 32 , 36 ] y muy poco sobre las especies interacciones [ 7 , 14 - 16 ]. En un ejemplo de esto último, el ruido del tráfico afecta negativamente murciélago ( Myotis Myotis ) la eficiencia de forrajeo al afectar su capacidad para localizar a sus presas por escuchar los sonidos generados por el movimiento presa [ 14 ]. Aquí, nuestros datos demuestran que la frecuencia de las interacciones entre especies puede cambiar sin un efecto directo de ruido en la propia interacción, lo que sugiere que la exposición al ruido puede provocar cambios en numerosas interacciones ecológicas y reverberan a través de las comunidades.
Los aumentos en las tasas de polinización estuvieron en línea con nuestra predicción basado en las respuestas positivas a ruido por A. alexandri , tanto en términos de la selección de sitios de anidación [ 7 ] y abundancias determinadas a partir de las encuestas [ 17 ]. Nuestro diseño experimental y el uso de flores artificiales eran ventajoso porque podíamos controlar la variación en la densidad y la distribución espacial de los recursos de néctar que pueden influir en los patrones de polinización [ 37 ]. Sin embargo, este enfoque nos impide determinar si el aumento de la polinización en los resultados de las zonas ruidosas en una mayor producción de semillas y frutos. Es probable por I. aggregata porque puede ser polen limitados en toda su gama de [ 38 - 40 ] y el cuajado se correlaciona fuertemente con los polinizadores (por ejemplo colibrí) abundancia [ 18 ]. Aumenta tanto, que dependen de ruido en A. Alexandri abundancias [ 7 , 17 ], junto con el aumento de las visitas a las flores artificiales en este estudio sugiere que I.aggregata plantas expuestas a niveles elevados de ruido pueden tener un mayor éxito reproductivo relativo a las personas en zonas tranquilas.
La remoción de semillas, depredación de semillas y plántulas de datos de reclutamiento eran coherentes entre sí y nuestras expectativas, lo que sugiere que el ruido tiene el potencial de afectar indirectamente la estructura del bosque. Es posible que el conjunto de especies eliminación de semillas puede variar en junio y julio, cuando se realizó el estudio de la encontrada en el otoño cuando las semillas son típicamente disponible. Sin embargo, todas las especies documentadas quitar las semillas son residentes todo el año y su abundancia relativa es poco probable que fluctúan entre el tratamiento y los centros de control durante todo el año.En cambio, es más probable que subestimar la magnitud de la diferencia en la calidad de la dispersión de semillas entre las zonas ruidosas y tranquilo por dos razones principales. En primer lugar, porque A. californicatípicamente provisión joven rico en proteínas presa animal [ 41 ], los individuos en nuestra área de estudio puede haber sido alimentándose principalmente de presas animales en lugar de P. edulis semillas durante nuestros experimentos. En segundo lugar, el uso de estaciones de semillas en el suelo no cuenta para la remoción de semillas de los conos en el dosel de otras importantes dispersores de semillas, tales como cyanocephalus Gymnorhinus (piñon jay), una especie que se produce en RCHMA, sino que también evita áreas ruidosas [ 7 , 42 ]. El grado en que estos factores contribuyen a reducir el reclutamiento de plántulas en áreas ruidosas se desconoce, pero ofrece una vía de investigación interesante para futuros estudios.
Aunque, A. californica y Peromyscus ratones tenían la mayor influencia en las tasas de remoción de semillas, no fue posible realizar el seguimiento del destino de las semillas individuales. Sin embargo, estas especies influyó en los patrones de depredación de semillas de una manera consistente con el conocimiento de cómo estas especies difieren como enlaces móviles para P.edulis dispersión de semillas y el establecimiento de plántulas. Evidencia de depredación de semillas fue menos frecuente en las estaciones de siembra visitados por A. californica , posiblemente reflejando su papel como dispersor importante de P. edulis semillas. Por ejemplo, uno A. californica individuo puede almacenar en caché hasta 6000 P. edulis semillas en lugares favorables para la germinación durante un otoño único [ 43 ]. Muchas semillas son reubicados y se consume, pero muchos van y no recuperado germinar [ 20 ]. Por el contrario, aunque Peromyscus ratones podría funcionar como dispersores condicional bajo ciertas circunstancias [ 44 , 45], aquí su presencia en una estación de semilla fue un fuerte predictor de la depredación de semillas, lo que refleja su papel principal como depredadores de semillas [ 20 ]. La investigación anterior usando recintos experimentales para estudiar el comportamiento de almacenamiento en caché en el campo apoya nuestros resultados [ 29 , 44 ]. Peromyscusratones consumen una gran proporción (aproximadamente 40%) de semillas encontradas y típicamente caché muchas semillas encontradas que no son consumidos inmediatamente [ 44 ] . Sin embargo, las semillas almacenadas en caché se han recuperado y se come (más del 80%) junto con las semillas almacenadas en caché por otros individuos o especies [ 29]. Por lo tanto, la reducción de la densidad de plántulas en áreas ruidosas podría explicarse no sólo por menos semillas que entran en el banco de semillas como resultado de la reducción de las densidades de importantes dispersores de semillas de aves que miles de caché muchas de las semillas, sino porque las semillas presentes en el banco de semillas de elevada experiencia las tasas de depredación por hurto caché asociada con cargo al ruido aumenta en Peromyscus ratones.
A pesar de la concordancia entre nuestros resultados y la literatura con respecto a los roles de A. californica y Peromyscus ratones con P. edulisdispersión de semillas y depredación, la mortalidad de plántulas causado por los depredadores de plántulas clave, como Odocoileus hemionus (venado bura) y Cervus canadensis (alce), podría explicar la mayor densidad de plántulas en relativa calma para áreas ruidosas. Sin embargo, los ungulados como C. canadensis parecen evitar áreas expuestas a ruido de alto volumen de tráfico [ 46 ], lo que sugiere que la mortalidad de plántulas debido a la navegación ungulados debe ser mayor en las zonas con menos ruido y que conduce a un patrón contrario de lo que se observó. Aún se necesita son estudios confirmatorios que realizan un seguimiento del destino de las semillas almacenadas en caché de documentos y patrones de depredación de plántulas en áreas ruidosas y tranquilo.
A pesar de las consecuencias posteriores de especies específicas de respuesta a la exposición al ruido, las razones mecánicas para cada especie, las respuestas aún no están claras. Aphelocoma californica pueden evitar zonas ruidosas porque el ruido puede enmascarar su comunicación vocal. Las aves más grandes con vocalizaciones de baja frecuencia son más sensibles al ruido que las especies más pequeñas con vocalizaciones de alta frecuencia debido a que sus vocalizaciones se superponen bajas frecuencias donde el ruido tiene más energía acústica [ 17 ]. Aphelocoma californica es también el principal depredador de nidos en el área de estudio [ 7 , 16 ], y es posible que las señales de ruido que enmascara utiliza para localizar a sus presas en nidos (por ejemplo, conferencias y polluelos de los padres). También es posible que estas formas de interferencia acústica puede conducir a niveles elevados de estrés que podrían influir en los patrones de uso del hábitat [ 13 ], pero la investigación sobre este posible vínculo hoy no existe.
En contraste con el ruido efecto directo puede tener en A. californicacomunicación y las respuestas positivas a la alimentación, el ruido por A.alexandri y Peromyscus ratones probablemente reflejan las respuestas indirectas al ruido. Áreas ruidosas pueden representar refugios contra los depredadores y competidores clave que suelen evitar zonas ruidosas, como los arrendajos. Por ejemplo, A. alexandri preferentemente pueden instalarse en áreas ruidosas en respuesta a las señales que indican una menor presión de depredación de nidos de A. californica . De manera similar,Peromyscus ratones poblaciones puede aumentar en las zonas ruidosas no sólo por reducción de la competencia con A. californica y otro para los arrendajos clave de forrajeo recursos, pero también en respuesta a la depredación reducida por los depredadores nocturnos acústicos que pueden evitar el ruido [ 14 ], como los búhos.
Ese ruido puede alterar los patrones de reclutamiento de plántulas agrega información importante a nuestro trabajo anterior donde encontramos ni P.edulis densidad de árboles, ni 12 características del hábitat difirió entre otros sitios de tratamiento y de control [ 7 ]. Esto, sin embargo, puede estar cambiando lentamente. Reducción del P. edulis reclutamiento de plántulas en áreas ruidosas eventualmente puede traducirse en un menor número de árboles maduros, sin embargo por P. edulis crece lentamente y tiene largos tiempos de generación [ 47 ], estos cambios iniciales en la estructura de la masa podría haber pasado desapercibido durante décadas. Tales cambios a largo plazo pueden tener implicaciones importantes para la comunidad forestal en su conjunto mediante la prórroga de las consecuencias negativas de la exposición al ruido. Es decir, el ruido no sólo puede dar lugar a grandes disminuciones en la diversidad durante la exposición al provocar el abandono del sitio o la reducción de las densidades de muchas especies [ 7 , 10 ], pero la diversidad pueda sufrir mucho después de que las fuentes de ruido se han ido porque menos P. edulis árboles proporcionan un hábitat menos crítica para los cientos de especies que dependen de ellos para su supervivencia [ 48 ].
Estos experimentos separados destacar que la contaminación acústica es una fuerza fuerte del medio ambiente que pueden alterar los procesos ecológicos fundamentales y servicios. Hace una década, Forman [ 4 ] estima que aproximadamente una quinta parte de la superficie terrestre en los Estados Unidos se ve afectado por el ruido del tráfico, a la medida geográfica real de la exposición al ruido es sin duda mayor cuando se consideran otras fuentes. Además, esta huella espacial del ruido, el paisaje sonoro antropogénico, no hará sino aumentar ya que las fuentes de contaminación acústica están creciendo a un ritmo más rápido que la población humana [ 3 ]. Estos datos sugieren que los sonidos antropogénicos tienen o van a abarcar casi todos los tipos de hábitats terrestres, que podría afectar las interacciones de innumerables especies, tanto directa como indirectamente. Es muy importante que identifiquemos que otras especies únicas funcionalmente abandonar o preferentemente instalarse en otras zonas ruidosas de todo el mundo. La detección temprana de distribución de las especies alteradas y las resultantes interrumpidas o mejorar los servicios ecológicos será la clave para entender la trayectoria de las muchas poblaciones y comunidades que por fuera parecen persistir a pesar de nuestra rumble industrial.
Agradecimientos
Este estudio se realizó en cumplimiento de la Universidad de Colorado Cuidado de Animales y el empleo Comisión.
Agradecemos a nuestros muchos asistentes de investigación, además de C. Quintero, J. Paritsis y S. Wagner, por su ayuda con los experimentos de polinización. También damos las gracias CA Botero, McClain CR y los árbitros anónimos por sus útiles sugerencias y comentarios sobre versiones anteriores de este manuscrito. Este trabajo fue apoyado principalmente por la NSF DDIG (n º 0910092 IOS), Estados Unidos Oficina de Administración de Tierras, ConocoPhillips, Energía Williams y U. Colorado del Departamento de Ecología y Biología Evolutiva. CDF fue apoyada por el Centro Nacional de Síntesis Evolutiva (NESCent; NSF EF-0905606).
· Recibido el 31 de enero 2012.
· Aceptado 28 de febrero 2012.
· Esta revista es © 2012 La Real Sociedad
Referencias
1. ↵
1. Ellis E. C.
2011 transformación antropogénica de la biosfera terrestre. Phil. Trans. R. Soc. A 369 , 1010 - 1035 . ( doi: 10.1098/rsta.2010.0331 )
2. ↵
1. Ellis E. C. ,
2. Ramankutty N.
2008 Poner a la gente en el mapa: biomas antropogénicos del mundo . Front. Ecol. . Medio 6 , 439 - 447 . ( doi: 10.1890/070062 )
3. ↵
1. Barber J. R. ,
2. Crooks K. R. ,
3. Fristrup K. M.
2010 Los costos de la exposición crónica al ruido para los organismos terrestres . Tendencias Ecol. Evol. 25 , 180 - 189 . ( doi: 10.1016/j.tree.2009.08.002 )
4. ↵
1. Forman R. TT
2000 Estimación de la zona afectada por el sistema ecológicamente carretera en los Estados Unidos . Conserv. . Biol. 14 ,31 - 35 . ( doi: 10.1046/j.1523-1739.2000.99299.x )
5. ↵
1. Francis C. D. ,
2. Paritsis J. ,
3. Ortega C. P. ,
4. Cruz A.
2011 Paisaje patrones de uso del hábitat aviar y el éxito de anidación se ven afectados por el gas crónico y el ruido del compresor . Ecol Paisaje. 26 , 1269 - 1280 . (doi: 10.1007/S10980-011-9609-Z )
6. ↵
1. Francis C. D. ,
2. Ortega C. P. ,
3. Cruz A.
2011 cambio de frecuencia vocal refleja las diferentes respuestas al ruido antropogénico en dos suboscine papamoscas tirano . Proc. R. Soc. B 278 , 2025 - 2031 . (doi: 10.1098/rspb.2010.1847 )
Abstract / FREE Texto completo
7. ↵
1. Francis C. D. ,
2. Ortega C. P. ,
3. Cruz A.
2009 La contaminación acústica cambia de comunidades de aves e interacciones entre especies . Curr. . Biol. 19 , 1415 - 1419 . ( doi: 10.1016/j.cub.2009.06.052 )
8. ↵
1. Francis C. D. ,
2. Ortega C. P. ,
3. Cruz A.
2011 Las diferentes respuestas de comportamiento a ruido antropogénico por dos aves paseriformes estrechamente relacionados . biol. Lett. 7 , 850 - 852 . ( doi: 10.1098/rsbl.2011.0359 )
Abstract / FREE Texto completo
1. Halfwerk W. ,
2. Slabbekoorn H.
2009 Un mecanismo conductual que explica el ruido dependiente de la frecuencia en el uso urbano canto de los pájaros . Anim. Behav. 78 , 1301 - 1307 . ( doi: 10.1016/j.anbehav.2009.09.015 )
10. ↵
1. Bayne E. M. ,
2. Habib L. ,
3. Boutin S.
2008 Impactos del ruido antropogénico crónico de la actividad del sector energético en la abundancia de aves canoras en el bosque boreal . conserv. . Biol. 22 ,1186 - 1193 . ( doi: 10.1111/j.1523-1739.2008.00973.x )
11. ↵
1. Slabbekoorn H. ,
2. Ripmeester E. AP
2008 Birdsong y el ruido antropogénico: implicaciones y aplicaciones para la conservación . Mol. . Ecol 17 , 72 - 83 . ( doi: 10.1111/j.1365-294X.2007.03487.x )
1. Patricelli G. L. ,
2. Blickley J. L.
2006 comunicación aviar en ruido urbano: causas y consecuencias del ajuste vocal . Auk 123 , 639 - 649 . ( doi: 10.1642/0004-8038 (2006) 123 [639: ACIUNC] 2.0.CO; 2 )
13. ↵
1. Kight C. R. ,
2. Envuelva J. P.
2011 ¿Cómo y por qué los animales impactos ambientales de ruido: una revisión integradora y mecanicista .Ecol. Lett. 14 , 1052 - 1061 . ( doi: 10.1111/J.1461-0248.2011.01664.X )
14. ↵
1. Siemers B. M. ,
2. Schaub A.
2011 La caza en la carretera: el ruido del tráfico reduce la eficiencia de forrajeo de los predadores acústicas . Proc.R. Soc. B 278 , 1646 - 1652 . ( doi: 10.1098/rspb.2010.2262 )
Abstract / FREE Texto completo
1. Schaub A. ,
2. Ostwald J. ,
3. Siemers B. M.
2008 murciélagos forraje evitar el ruido . J. Exp. . Biol. 211 , 3174 - 3180 . ( doi: 10.1242/jeb.022863 )
Abstract / FREE Texto completo
16. ↵
1. Francis C. D. ,
2. Ortega C. P. ,
3. Kennedy R. I. ,
4. Nylander P. J.
En prensa.Son depredadores de nidos ausente de las zonas ruidosas o incapaces de localizar nidos ? Ornithol. Monogr .
17. ↵
1. Francis C. D. ,
2. Ortega C. P. ,
3. Cruz A.
2011 Filtros de ruido la contaminación de aves comunidades basadas en la frecuencia vocal .PLoS ONE 6 , e27052 . ( doi: 10.1371/journal.pone.0027052 )
18. ↵
1. Paige K. N. ,
2. Whitham T. G.
1987 flexibles rasgos de historia de vida: cambios de color escarlata gilia en respuesta a la abundancia de polinizadores . Ecología 68 , 1691 - 1695 . ( doi: 10.2307/1939861 )
1. Vander Wall S. B. ,
2. Balda R. P.
1981 Ecología y evolución de la conducta alimentaria de almacenamiento en caché de córvidos coníferas de semillas . Z. Tierpsychol. 56 , 217 - 242 . ( doi: 10.1111/j.1439-0310.1981.tb01298.x )
20. ↵
1. Cámaras J. C. ,
2. Vander Wall S. B. ,
3. Schupp E. W.
1999 de semillas y plántulas de piñón ecología y especies de enebro en los bosques enanos del oeste de Norteamérica . Bot. Rev. 65 , 1 - 38 . ( doi: 10.1007/BF02856556 )
21. ↵
1. Hurly T. A.
1996 La memoria espacial en colibríes rufous: la memoria de los sitios premiados y no premiados, . Anim. Behav. 51 , 177 - 183 . (doi: 10.1006/anbe.1996.0015 )
22. ↵
1. Hurly T. A. ,
2. Franz S. ,
3. Healy S. D.
2010 No colibríes rufo ( Selasphorus rufus ) utilizan balizas visuales? Anim. Cogn. 13 , 377 - 383 . ( doi: 10.1007/s10071-009-0280-6 )
23. ↵
1. Baum K. A. ,
2. Subvención W. E.
2001 Hummingbird comportamiento de forrajeo en los tipos de conexión diferentes: simulación de estrategias alternativas . Ecol. Modelo. 137 , 201 - 209 . ( doi: 10.1016/S0304-3800 (00) 00.436-1 )
24. ↵
1. Pleasants J. M.
1983 los patrones de producción de néctar enIpomopsis aggregata (Polemoniaceae) . Soy. J. Bot. 70 , 1468 - 1475 .
25. ↵
1. Waser N. M. ,
2. McRobert J. A.
1998 colibrí alimentándose de parches experimentales de flores: evidencia de débil aversión al riesgo . J. Biol. aviar. 29 , 305 - 313 .
26. ↵
R Core Development Team 2010 R: un lenguaje y un entorno para computación estadística . Viena, Austria : R Foundation for Statistical Computing .
27. ↵
1. Dudash M. R.
1991 Planta efectos de tamaño en función femenina y masculina en hermafrodita Sabatia angularis (Gentianaceae) . Ecología72 , 1004 - 1012 . ( doi: 10.2307/1940600 )
28. ↵
1. Kearns C. A. ,
2. Inouye D. W.
1993 Técnicas para la polinización de los biólogos . Niwot, CO : University Press of Colorado .
29. ↵
1. Vander Wall S. B.
2000 La influencia de las condiciones ambientales sobre la recuperación de la memoria caché y caché de hurto por el amarillo ardillas del pino ( Tamias amoenus ) y ratones ciervo (Peromyscus maniculatus ) . Behav. . Ecol 11 , 544 - 549 . ( doi: 10.1093/beheco/11.5.544 )
Abstract / FREE Texto completo
30. ↵
1. Harrington M. G.
1987 Características de un año de edad, las plántulas de piñón naturales. Nota de Investigación RM-477 . Fort Collins, CO : USDA Forest Service, Rocky Mountain Research Station .
31. ↵
1. Segura G. ,
2. Snook L. C.
1992 Soporte dinámica y patrones de regeneración de un bosque de pinos piñoneros en el este central de México . Ecol Bosque. Manag. 47 , 175 - 194 . ( doi: 10.1016/0378-1127 (92) 90273-C )
32. ↵
1. Halfwerk W. ,
2. Holleman L. JM ,
3. Lessells C. M. ,
4. Slabbekoorn H.
2011Impacto negativo del ruido del tráfico en el éxito reproductivo aviar . J.Appl. . Ecol 48 , 210 - 219 . ( doi: 10.1111/j.1365-2664.2010.01914.x )
33. ↵
1. Summers P. D. ,
2. Cunnington G. M. ,
3. Fahrig L.
2011 ¿Los efectos negativos de las carreteras en las aves reproductoras causadas por el ruido del tráfico? J. Appl. . Ecol 48 , 1527 - 1534 . ( doi: 10.1111/j.1365-2664.2011.02041.x )
34. ↵
1. Bruto K. ,
2. Pasinelli G. ,
3. Kunc H. P.
2010 plasticidad conductual permite ajuste a corto plazo a un nuevo entorno . Soy. Nat. 176 , 456 - 464 . (doi: 10.1086/655428 )
35. ↵
1. Halfwerk W. ,
2. Bot S. ,
3. Buikx J. ,
4. van der Velde M. ,
5. Komdeur J.,
6. Ten Cate C. ,
7. Slabbekoorn H.
2011 de baja frecuencia canciones perder su potencia en condiciones urbanas ruidosas . Proc. Natl Acad. Sci.EE.UU. 108 , 549 14 - 554 14 . ( doi: 10.1073/pnas.1109091108 )
Abstract / FREE Texto completo
36. ↵
1. Habib L. ,
2. Bayne E. M. ,
3. Boutin S.
2007 ruido industrial crónica afecta a la estructura de apareamiento éxito y la edad de horneros Seiurus aurocapilla . J. Appl. . Ecol 44 , 176 - 184 . ( doi: 10.1111/j.1365-2664.2006.01234.x )
37. ↵
1. Feinsinger P. ,
2. Iii H. MT ,
3. Joven B. E.
1991 ¿Es tropicales plantas polinizadas por aves presentan interacciones dependientes de la densidad? Los experimentos de campo . Ecología 72 , 1953 - 1963 . (doi: 10.2307/1941550 )
38. ↵
1. Hainsworth F. R. ,
2. Lobo L. L. ,
3. Mercier T.
1985 limitación de polen de una especie monocárpica, Ipomopsis aggregata . J. . Ecol 73 , 263 - 270 .
1. Campbell D. R. ,
2. Halama K. J.
1993 Recursos y polen limitación s para la producción de semillas de por vida en una población natural de la planta. Ecología 74 , 1043 - 1051 . ( doi: 10.2307/1940474 )
40. ↵
1. Juenger T. ,
2. Bergelson J.
1997 El polen y la limitación de los recursos de la indemnización a la herbivoría en escarlata gilia, Ipomopsis aggregata . Ecología 78 , 1684 - 1695 . ( doi: 10.1890/0012-9658 (1997) 078 [1684: PARLOC] 2.0.CO; 2 )
41. ↵
1. Curry R. L. ,
2. Peterson A. T. ,
3. Langen T. A.
2002 Western fregar-jay (Aphelocoma californica ), no. 712 . En las aves de América del Norte (ed.Poole A. ), pp 1 - 36 . Ithaca, NY : Cornell Laboratorio de Ornitología .
42. ↵
1. Ortega C. P. ,
2. Francis C. D.
En prensa. Efectos del ruido del compresor de gas y en la capacidad para detectar aves en las encuestas en el noroeste de Nuevo México . Ornithol. Monogr .
43. ↵
1. Balda R. P.
1987 impacto aviar en piñón-junípero bosque . En Proc.piñón-junípero Conf, Informe General Técnico INT-215. (ed. Everett RL ), pp 526 - 533 . Ogden, UT : Bosque Intermountain y Estación Experimental Range .
44. ↵
1. Pearson K. M. ,
2. Theimer T. C.
2004 Semillas de almacenamiento en caché las respuestas a sustrato y cubierta de roca por dos Peromyscusespecies: implicaciones para el establecimiento de pinos piñoneros .Oecologia 141 , 76 - 83 . ( doi: 10.1007/s00442-004-1638-8 )
45. ↵
1. Theimer T. C.
2005 dispersores de semillas roedores como mutualistas condicionales . En el destino de semillas: establecimiento depredación, dispersión y plántulas (eds Olvídate P.-M. , Lambert JE ,Hulme PE , Vander Wall SB ), pp 283 - 296 . Cambridge, MA : CABI Press.
46. ↵
1. Gagnon J. W. ,
2. Theimer T. C. ,
3. Boe S. ,
4. Dodd N. L. ,
5. Schweinsburg R. E.
2007 El volumen de tráfico altera la distribución de los alces y los cruces de carreteras en Arizona . J. Wildl. Administrar. 71 , 2318 - 2323 . ( doi: 10.2193/2006-224 )
47. ↵
1. Floyd M. L. ,
2. Colyer M. ,
3. Hanna D. D. ,
4. Romme W. H.
2003 árboles viejos Gnarly: características del dosel de edad madura piñón-junípero . Enantiguas piñón-junípero (ed. Floyd ML ), páginas 11 - 29 . Boulder, CO :Universidad de Colorado Press .
48. ↵
1. Mueller R. C. ,
2. Scudder C. M. ,
3. Porter M. E. ,
4. Trotter R. T. ,
5. Gehring C. A.,
6. Whitham T. G.
2005 Diferencial mortalidad de los árboles en respuesta a la grave sequía: evidencia de los cambios de vegetación a largo plazo. J. . Ecol 93 , 1085 - 1093 . ( doi: 10.1111/j.1365-2745.2005.01042.x )
Blog: Http://apiculturauruguay.blogspot.com/



No hay comentarios:
Publicar un comentario